Practical medical trials have confirmed the maximum effect of Levitra, whith a positive effect being observed in 99 of 100 patients wellbutrin xl uk Our internet store sells medicines produced in Indian pharmaceutical factories under licensed agreement.
Ophtalmo.ulg.ac.be
Estrogens Reduce the Expression of YKL-40 in the Retina: Implications for Eye and Joint Diseases Jean-Marie Rakic,1 Vincent Lambert,2 Manuel Deprez,3 Jean-Michel Foidart,2 AgnePURPOSE. To identify modifications in the gene expression pro-
mer’s disease.9 Menopause coincides also with the appearance
file of the ocular posterior segment in ovariectomized (OVX)
or worsening of many common arthritic conditions10 that are
mice with and without substitutive estradiol therapy and to
associated with increased levels of proinflammatory cytokines,
select differentially expressed genes that could be relevant to
such as interleukin (IL)-1, IL-6, and tumor necrosis factor-␣.11
the natural history of human age-related macular degeneration
Epidemiologic studies reported conflicting results on the pro-
tective effect of hormone replacement therapy (HRT) in the
development of the neovascular form of AMD.12,13 Others
ETHODS. Chorioretinal tissues from two groups of 25 treated
and untreated OVX mice were analyzed by using cDNA array
studies, however, failed to demonstrate a significant gender
technology. The expression level of selected genes was con-
difference in the frequency of the disease.14 A large clinical
firmed in triplicate by RT-PCR and related to the estrogenic
trial (Women’s Health Initiative Sight Exam Study) investigating
status of the animals. Expression of the YKL-40 gene was
whether HRT may influence the development and the course
further investigated in intact or diseased human retinas and in
of macular degeneration is under way.
a murine model of experimental choroidal neovascularization
The mechanisms by which 17--estradiol (E2) protects
(CNV), using laser pressure catapulting.
against AMD are not known. A protective effect has beendemonstrated in the ischemia–reperfusion injury model in rat
RESULTS. Of the approximately 10,000 genes screened, only
retina.15 E2 enhances the expression of vascular endothelial
YKL-40 expression was significantly downregulated by 17--
growth factor (VEGF) and its receptor (VEGFR-2) in bovine
estradiol. YKL-40 was expressed in intact human neural retina
retinal endothelial cells in vitro,16 but this effect has recently
and in the RPE. The expression of YKL-40 was upregulated in
been shown to depend on oxygen status in a murine model of
experimental CNV and in neovascular membranes extracted
retinopathy of prematurity.17 Ovariectomy has been suggested
from patients affected by the exudative form of AMD.
to disturb Bruch’s membrane turnover in mice fed with a
CONCLUSIONS. These observations indicate that YKL-40 expres-
high-fat diet (Alexandridou A, Elliot S, Espinosa D, Hernandez
sion in the retina is modulated by serum levels of estradiol. This
E, Csaky KG, Cousins SW, ARVO Abstract 1200, 2001).
protein could be relevant to the development of AMD and is
In the present study, we used cDNA microarray technology
also a new mediator to take into account when evaluating the
to profile gene expression changes (ϳ10,000 screened genes)
broad consequences of hormonal replacement therapy. (Invest
in intact posterior segments of ovariectomized (OVX) mice
Ophthalmol Vis Sci. 2003;44:1740 –1746) DOI:10.1167/iovs.02-
treated or not with E2 pellets. In a second confirmation phase,
the expression of selected genes was investigated by quantita-tive RT-PCR on individual mice retinas and correlated with the
Thepresenceofbothtypesofestrogenreceptors(ER)␣and estrogen status. E2 supplementation significantly reduced the
 has recently been demonstrated in the retina, suggesting
expression of one gene, YKL-40 (also known as human carti-
a role for an intracellular receptor-mediated effect of estrogens
lage glycoprotein 39, HCgp39). YKL-40 is a mammalian glyco-
in retinal biology.1,2 The protective role of estradiol in the
protein member of a family of 18 glysolyl hydrolases related in
development of cataract has been convincingly established on
sequence to chitinases but without chitinase activity.18,19 Its
experimental3,4 and epidemiologic grounds.5 However, the
function is unknown, but it is thought to be involved in tissue
precise effects of estrogens on the retina remain largely unex-
remodeling, acting as a growth factor for connective tissue
cells20 and as a potent migration factor for endothelial cells.21
Increasing experimental evidence suggests that estrogens
Most of the studies regarding YKL-40 have focused on rheuma-
are neuroprotective6 – 8 and that estrogen replacement therapy
toid arthritis (RA), because this protein is an articular autoan-
may contribute to the prevention or delay the onset of Alzhei-
tigen in mice22 (with pathologic effects similar to those withimmunization by collagen type II), and its serum levels areclinically correlated with the severity of the disease.23,24 Be-
From the 1Department of Ophthalmology, University Hospital,
cause this glycoprotein has not been previously described in
`ge, Belgium; the Laboratories of 2Tumor and Develop-
the retina, we studied its expression in human normal retina, in
ment Biology and 3Neuropathology, University of Lie
neovascular choroidal membranes extracted from patients
with AMD, and in a murine model of experimental choroidal
Supported by grants from Les Amis des Aveugles, Ghlin, Belgium.
CM is a research associate and AN a senior research associate from theNational Fund for Scientific Research (FNRS), Belgium.
Submitted for publication August 1, 2002; revised August 29 and
September 19, 2002; accepted September 25, 2002.
Disclosure: J.-M. Rakic, None; V. Lambert, None; M. Deprez,
None; J.-M. Foidart, None; A. Noe ¨l, None; C. Munaut, None Ovariectomized (OVX) Mice
The publication costs of this article were defrayed in part by page
OVX mice (ovariectomy was performed at 6 weeks of age) were
charge payment. This article must therefore be marked “advertise-
purchased from Charles River France (Lyon, France). In the group (n ϭ
ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
25) receiving substitutive hormonal therapy, estrogens were adminis-
Corresponding author: Jean-Marie Rakic, Polyclinique d’Ophtal-
mologie, Centre Hospitalier Universitaire, Sart-Tilman, B-4000 Lie
tered through 3-mm subcutaneous pellets containing 1.7 mg E2 (Inno-
vative Research of America, Sarasota, FL). In the control group, OVX
Investigative Ophthalmology & Visual Science, April 2003, Vol. 44, No. 4
Copyright Association for Research in Vision and Ophthalmology
Effect of Estrogens on Retinal YKL-40 TABLE 1. Oligonucleotide Sequences Size of PCR Product (Accession No.) Position Oligonucleotide Sequence (5–3) Cycles (n)
m and h, mouse and human, respectively.
mice (n ϭ 25) received implants of a saline-containing pellet. E2 pellets
with exudative AMD (three men, five women; mean age, 77 years;
provide continuous release, allowing for a constant level of circulating
range, 72– 83). These patients were not eligible for conventional laser
hormone. The selected pellet delivered 28 g/d of E2, providing serum
photodynamic therapy because of the presence of occult new vessels
concentrations exceeding the usual physiological concentrations in
or submacular bleeding, or, in the case of one patient, because of a
female mice (ϳ100 pg/mL) but lower than high-dose estrogen treat-
recurrence of the disease. The methods conformed to the Declaration
ment (e.g., anti-tumor therapy).25 Serum levels of E2 were monitored
of Helsinki for research involving human subjects. The specimens were
at the end of the study period by radioimmunoassay, according to the
snap frozen in liquid nitrogen and stored at Ϫ80°C. Ten human male
manufacturer’s instructions (Immunotech; Westbrook, ME), as previ-
and female donor eyes were also obtained from the Cornea Bank,
ously described.2 After 30 days, mice were killed, and the ocular
`ge, Belgium, as a source of intact neural retina and
posterior segments (neural retina and RPE-choroid complex) were
RPE-choroid. After removal of the anterior segment structures, 5-mm
removed and frozen in liquid nitrogen. In the treated group, the
diameter punches were made in the macular region and in the periph-
efficiency of HRT was also controlled by means of uterine weight
eral retina (to estimate differential expression between the two re-
(approximate fivefold increase in the E2 treated group) and histologic
gions), frozen in liquid nitrogen, and stored at Ϫ80°C.
evaluation of the endometrium (not shown). Three mice had to beremoved from the study group because of defective pellet implanta-
Laser Pressure Catapulting
tion. For experiments using RT-PCR, a second set of 20 OVX mice (10
For laser pressure catapulting (LPC), experimental CNV was induced
in each group) was treated similarly (2 mice were not analyzed in the
by laser in C57BL mice (n ϭ 12), as previously described,26 to evaluate
E2-treated group because of bad pellet implantation). Animal experi-ments were performed in compliance with the ARVO Statement forthe Use of Animals in Ophthalmic and Vision Research. cDNA Array
Total RNA was isolated from posterior segments (retina and RPE-choroid complex) from OVX control and OVX-E2–treated mice byusing an RNA isolation protocol (TRIzol; Sigma, Belgium), as recom-mended by the manufacturer. The frozen tissues were first pulverizedwith a dismembrator (B. Braun Biotech International, GmBH, Melsun-gen, Germany) and homogenized in the extraction reagent. RNA qual-ity was controlled on 1% agarose gel. RNA samples from each groupwere pooled and then processed by Incyte Genomics (St. Louis, MO)and hybridized to a cDNA array (Mouse UniGene1; Incyte Genomics,Inc.). This array is designed to provide a broad view of the mousegenome and includes targets for 9596 elements with 8985 uniqueannotated genes and expressed sequence tag (EST) clusters (list avail-able at http://www.incyte.com/expression/easy_to_spot/catalog/txt_files/mouse_unigene1_gene_list.txt). Gene expression levels weremeasured and displayed using the manufacturer’s software (GEMTools2.5; Incyte Genomics). Human Intact and Pathologic Retinas
Eight consecutive submacular CNV specimens were surgically re-
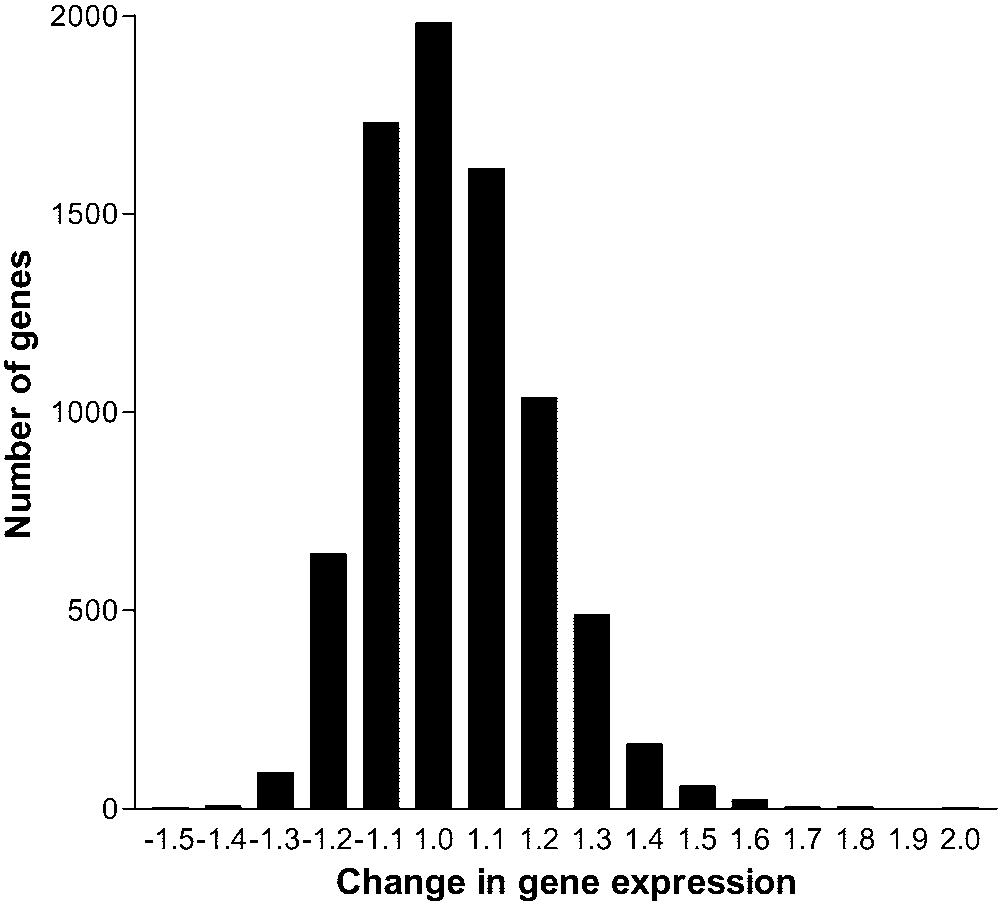
FIGURE 1.
Frequency of genes exhibiting different levels of expres-
moved (performed by Carel Claes, MD, A.Z Middelheim, Antwerpen,
sion (in multiples of change) in OVX mice supplemented or not with
Belgium) during 360° macular translocations performed on patients
E2 pellets (increased expressions are shown as negative values). Rakic et al. TABLE 2. Gene Expression in the Absence of E2 Treatment Accession Effects of E2 on Chorioretinal mRNA Profile Gene Description
The mean E2 serum level was 15.08 Ϯ 4.2 pg/mL in OVX-
untreated mice and 537 Ϯ 79.7 pg/mL in E2-treated mice,
indicating that ovariectomy reduced endogenous circulating
E2 concentrations, whereas supplementation with 1.7-mg es-
tradiol pellets restored circulating concentrations of E2 mod-
erately over the physiological range (100 pg/mL).25 Only a
minority of the genes tested on the cDNA array showed a
noticeable change of their expression after 30 days of substi-
tutive E2 therapy. Less than 1% of tested genes displayed a
differential expression ratio of 1.5 or more, and most of them
appeared to be upregulated by E2 deficiency (Fig. 1). The
Insulin-like growth factor–binding protein 4
cDNA microarray expression data of selected genes are shown
in Table 2. The largest difference in expression (ratio ϭ 2) was
seen for the YKL-40 gene with lower expression in OVX
E2-treated mice. The influence of E2 supplementation on
Matrix ␥-carboxyglutamate (gla) protein
YKL-40 expression was confirmed by semiquantitative RT-PCR
(in triplicate and with two different couples of oligonucleo-
tides normalized to 28S signal) performed on a second set of
cDNA microarray expression data of selected genes analyzed on
RNA samples pooled from E2-treated (n ϭ 22) or E2-untreated (n ϭ 25)OVX mice. All expression changes are positive except for HIF-␣.
the spatial and temporal pattern of YKL-40 expression. The eyes wereenucleated at selected intervals after induction (days 3, 6, 14, and 40),embedded in mounting compound (Tissue Tek; Miles Laboratories,Naperville, IL), and frozen in liquid nitrogen. Serial frozen sections(n ϭ 8 –10) were mounted directly on 1.35-mm thin polyethylene foil(Palm, Wolfratshausen, Germany), which had been mounted on theglass slide using a robotic technique (Microbeam-Moment; Palm).27The robot microbeam (Palm) focused the laser (337 nm) on thespecimen, catapulting the selected area into the microfuge cap. Theentire subretinal CNV area (as shown in Fig. 4C) and an adjacent intactchorioretinal zone (control) were microdissected separately on frozensections (10 m thick). The specimens were covered with 100 L lysisbuffer, and total RNA isolation was performed with an RNA isolation kit(PureScript RNA; BioZym, Landgraaf, The Netherlands) according tothe manufacturer’s protocol. Total RNA was dissolved in a 10 L RNAhydration solution supplied by the manufacturer, and RT-PCR wasperformed. RT-PCR Analysis
The frozen tissues were pulverized in a dismembrator (B. Braun Bio-tech International), and total RNA was extracted with a kit (RNeasy;Qiagen, Paris, France), according to the manufacturer’s protocol. 28SrRNA was amplified with an aliquot of 10 ng total RNA (for tissues) or1 L total RNA (LPC samples), by using an RNA PCR kit (GeneAmpwith Thermostable rTth reverse transcriptase; Applied Biosystems,Foster City, CA) and two pairs of primers (Eurogentec, Lie
Oligonucleotide sequences are shown in Table 1. Reverse transcriptionwas performed at 70°C for 15 minutes followed by 2 minutes incuba-tion at 95°C for denaturation of RNA-DNA heteroduplexes. Amplifica-tion was performed in cycles of 15 seconds at 94°C, 20 seconds at60°C, and 10 seconds at 72°C. RT-PCR products were resolved on 2%agarose gels and analyzed by fluorescence imager (Fluor-S MultiImager;BioRad, Richmond, CA), after staining with ethidium bromide (FMCBioProducts, Philadelphia, PA). FIGURE 2.
Influence of E2 treatment on YKL-40 gene expression in
Statistics
OVX mice. (A) Representative example of YKL-40 mRNA expression in the eye of individual mice. (B) Relative levels of YKL-40 mRNA (den-
Gene expression levels in both groups were compared on computer by
sitometric quantification and normalization to the 28S signal) in the
t-test (Prism 3.0; GraphPad, San Diego, CA).
E2-treated (ϩE2) and untreated (ϪE2) group. Effect of Estrogens on Retinal YKL-40 FIGURE 3.
of the cDNA array results for selectedgenes (used as negative control). Transthyretin, retinol binding pro-tein (RBP), angiogenin, and VEGF-AmRNA expression levels (densito-metric quantification and normaliza-tion to 28S signal) in ocular posteriorsegments of individual mice. Therewas no statistically significant differ-ence between mean mRNA expres-sion in the E2-treated or untreatedOVX mice (P Ͼ 0.05, t-test).
animals and related to the presence or absence of E2 supple-
temporal pattern of YKL-40 expression, RT-PCR analysis was
mentation (Fig. 2). Several genes showing similar expression
applied to laser-induced murine neovascular choroidal mem-
levels in the E2-treated and untreated groups were assessed in
branes microdissected (Fig. 4C, left) by LPC at different time
parallel by RT-PCR and used as the negative control (Fig. 3).
end points (days 3– 40 after laser-induced CNV). When com-
Table 3 shows the relative expression (signal intensity) of
pared with intact adjacent regions, upregulation of YKL-40
several groups of genes potentially involved in chorioretinal
mRNA expression was evident in CNV lesions at all end points
diseases in the untreated and E2-treated group.
It is interesting to note the relatively high level of expres-
sion of CD59a (an inhibitor of complement activation28), in
view of the recent reports of complement involvement as a
ISCUSSION
drusen component29 and as a mediator of CNV (Bora PS, Sohn
DNA microarrays are powerful tools that allow for genome-
JH, Kang SG, Bora NS, Kaplan HJ, ARVO Abstract 1291, 2002).
wide gene expression profiling of cells or tissues.32 In this
Elevated expression of -crystallin in the retina has been re-
study, we used a cDNA array testing for approximately 10,000
ported previously, without explanation of its potential role.30
genes to identify modification of the gene expression profile of
Membrane-type matrix metalloproteinases (MT1-MMP) and tis-
pooled ocular posterior segments in ovariectomized mice res-
sue inhibitor of matrix metalloproteinase (TIMP)-3 also had a
cued or not by substitutive E2 therapy. In this experimental
relatively high level of expression. Angiopoietin-like 2 had the
setting, less than 1% of the genes tested showed noticeable
highest level of expression in the angiogenesis regulator
expression changes. Among them, the YKL-40 gene showed
group,31 whereas some members of the VEGF family were
the largest expression change, with lower values in OVX E2-
weakly expressed (placental growth factor) or absent (VEGF-B
treated mice. This observation was confirmed by semiquanti-
tative RT-PCR in a second set of experiments performed on
YKL-40 Expression Profile in Human and Mouse
individual mice tissues. YKL-40 gene expression was furtherstudied in normal and pathologic human retinas. Neovascular Membranes
YKL-40 expression has been reported in human RPE, with
YKL-40 expression was detected in normal human posterior
serial analysis of gene expression technology (SAGE).33 In the
segments from different donors (Fig. 4A). YKL-40 was detected
current study, YKL-40 was consistently expressed in normal
in ocular tissues from patients of different age and gender,
human eyes both in neural retina and in the RPE-choroid
regardless of the region of the sample (inside or outside the
complex. High concentrations of YKL-40 in serum correlates
macula, retina or RPE-choroid complex). YKL-40 mRNA was
with morbidity in different human diseases such as active
also detected in all CNV specimens obtained during surgery
rheumatoid arthritis and hepatic fibrosis, as well as with death
(Fig. 4B). Although sharing in common the unfeasibility of
of recurrent colorectal cancer.34 –36 YKL-40 is produced by
conventional treatment, these neovascular membranes had a
monocytes in the media of arteritic vessels, in inflamed syno-
documented different natural history (size of the lesion, dura-
vial membranes, and in atherosclerotic plaques, suggesting a
tion of symptoms, previous treatment). To evaluate more pre-
role for YKL-40 in tissue remodeling.37,38 In vitro, YKL-40 is
cisely the spatial (lesion versus intact adjacent regions) and
one of the most abundant proteins secreted by cultured chon-
Rakic et al. TABLE 3. Relative Expression of Selected Groups of Genes Relative Level of Expression Gene Description Accession No. Untreated E2-Treated
cDNA microarray relative expression data normalized to microarray, internal control signals, and
analyzed on RNA samples pooled from E2-treated (n ϭ 22) or E2-untreated (n ϭ 25) OVX mice. PEDF,(pigmented epithelium-derived growth factor).
drocytes.39 The regulation of YKL-40 expression, however, is
our results could also provide a putative explanation for the
largely unknown. In this study YKL-40 was significantly down-
protective effect of estrogens in rheumatoid arthritis, because
regulated by estrogen supplementation in the ocular posterior
YKL-40 has been proposed as a candidate autoantigen in this
segment of OVX mice. Furthermore, YKL-40 expression was
disease,23 and its serum levels are related to disease activity.25
upregulated both in pathologic human and experimental CNV,
It is of interest that, in our model of OVX mice, only a
although we cannot exclude bias caused by our selecting
limited number of genes were affected by estrogen therapy in
specimens from patients for whom standard laser therapy
the retina. This could be explained by the pooling of samples
could not be used. Taken together with the migratory proper-
for cDNA array, which selected only for strong and constant
ties of YKL-40 in endothelial cells,21 these data suggest a
differences of gene expression and minimized individual vari-
proangiogenic role of YKL-40 in the development of exudative
ations. In particular, the expression of cathepsin D, an aspartic
AMD and could at least partly verify a protective role for
protease highly expressed in human retinal pigment epithelial
estrogen replacement therapy.12,13 A direct effect of YKL-40
lysosomes40 with transcription classically regulated by estro-
on angiogenesis in other in vitro and in vivo models, such as
gens in breast cancer,41 was not influenced in vivo in the
corneal pellets or aortic rings, has yet to be evaluated. If a
murine retina. Experimental impairment of cathepsin D results
similar regulation were to be demonstrated in the synovium,
in accumulation of rod outer segment debris in the RPE and as
Effect of Estrogens on Retinal YKL-40 FIGURE 4.
Expression of YKL-40 mRNA in human retina and in CNV. (A) YKL-40 mRNA was present in human normal neural retina (R) and in the
RPE-choroid complex (Ch), both in the macular region (I) and in the periphery (O). Age of the donors is given in years with gender. (B) Representative example of YKL-40 mRNA expression in surgically extracted choroidal neovascular membranes of four patients with exudative AMD. Total RNA (10 ng) was subjected to RT-PCR. (C) LPC followed by RT-PCR analysis of YKL-40 expression (with pigmented epithelium-derived growth factor [PEDF] used as an additional control gene on microdissected new choroidal vessels growing in the subretinal space (CNV, delineated by dotted line) and on adjacent (control) intact chorioretinal tissue (R) from days 3 to 40. Multiple bands under YKL-40 or PEDF signals represent multimers of primers. Molecular weights are shown at left (expected size for 28S product is 212 bp; for PEDF, 256 bp; for human YKL-40, 112 bp; and for murine YKL-40 142 bp).
been suggested as a murine model of dry AMD.42 A similar
the human retina, in which these receptors are expressed at a
discrepancy has been reported for endometrial cells and attrib-
uted to differential tissue-dependent regulation.43 Individual
This study is obviously a preliminary phase in the under-
variations in the level of mRNA expression for a specific gene
standing of the potential influences of HRT on AMD, and it
are evident from the confirmation phase of our study, espe-
would be of great interest to evaluate by cDNA microarray
cially in the OVX untreated group. There was up to a 15-fold
HRT-treated and untreated patients, as well as to compare
modulation of VEGF-A expression between mice of the same
intact and AMD tissue specimens. Our data nevertheless iden-
group. This could be explained by variations in very low level
tify for the first time the YKL-40 gene as a potential mediator of
E2 concentrations (picomolar range), which have been dem-
estrogens effects both in the normal eye and during the devel-
onstrated to enhance gene expression in different models.44 It
is also known that for some genes, the mechanism of regula-tion by estrogens is posttranscriptional rather than at the tran-
References
scriptional level. This was indeed demonstrated for hepaticapolipoprotein E.45 Finally, because the levels of ER-␣ and -
1. Ogueta SB, Schwartz SD, Yamashita CK, Farber DB. Estrogen re-
mRNA were weak in murine posterior segment, we cannot
ceptor in the human eye: influence of gender and age on gene
exclude the possibility that HRT could modify other genes in
expression. Invest Ophthalmol Vis Sci. 1999;40:1906 –1911. Rakic et al.
2. Munaut C, Lambert V, Noel A, et al. Presence of oestrogen receptor
25. Bourassa PAK, Milos PM, Gaynor BJ, Breslow JL, Aiello RJ. Estrogen
type beta in human retina. Br J Ophthalmol. 2001;85:877– 882.
reduces atherosclerotic lesion development in apolipoprotein E-
3. Bigsby RM, Cardenas H, Caperell-Grant A, Grubbs CJ. Protective
deficient mice. Proc Natl Acad Sci USA. 1996;93:10022–10027.
effects of estrogen in a rat model of age-related cataracts. Proc Natl
26. Lambert V, Munaut C, Noel A, et al. Influence of plasminogen
Acad Sci USA. 1999;96:9328 –9332.
activator inhibitor type I on choroidal neovascularization. FASEB J.
4. Davis VL, Chan CC, Schoen TJ, Couse JF, Chader GJ, Korach KS. An
estrogen receptor repressor induces cataract formation in trans-
27. Bohm M, Wieland I, Schutze K, Rubben H. Microbeam MOMeNT:
genic mice. Proc Natl Acad Sci USA. 2002;99:9427–9432.
non-contact laser microdissection of membrane-mounted native
5. Benitez del Castillo JM, del Rio T, Garcia-Sanchez J. Effects of
tissue. Am J Pathol. 1997;151:63– 67.
estrogen use on lens transmittance in postmenopausal women.
28. Meri S, Morgan BP, Davies A, et al. Human protectin (CD59), an
Ophthalmology. 1997;104:970 –973.
18,000 –20,000 MW complement lysis restricting factor, inhibits
6. Behl C. Oestrogen as a neuroprotective hormone. Nat Rev Neu-
C5b-8 catalysed insertion of C9 into lipid bilayers. Immunology.
7. Behl C, Widmann M, Trapp T, Holsboer F. 17-beta Estradiol pro-
29. Mullins RF, Russell SR, Anderson DH, Hageman GS. Drusen asso-
tects neurons from oxidative stress-induced cell death in vitro.
ciated with aging and age-related macular degeneration contain
Biochem Biophys Res Commun. 1995;216:473– 482.
proteins common to extracellular deposits associated with athero-
8. Kadish I, Van Groen T. Low levels of estrogen significantly dimin-
sclerosis, elastosis, amyloidosis, and dense deposit disease. FASEB
ish axonal sprouting after entorhinal cortex lesions in the mouse. J Neurosci. 2002;22:4095– 4102.
9. Tang MX, Jacobs D, Stern Y, et al. Effect of oestrogen during
30. Magabo KS, Horwitz J, Piatigorsky J, Kantorow M. Expression of
menopause on risk and age at onset of Alzheimer’s disease. Lancet.
betaB(2)-crystallin mRNA and protein in retina, brain, and testis. Invest Ophthalmol Vis Sci. 2000;41:3056 –3060.
10. Wluka AE, Cicuttini FM, Spector TD. Menopause, oestrogens and
31. Xu ZP, Tsuji T, Riordan JF, Hu GF. The nuclear function of angio-
arthritis. Maturitas. 2000;35:183–199.
genin in endothelial cells is related to rRNA production. Biochem
11. Pfeilschifter J, Koditz R, Pfohl M, Schatz H. Changes in proinflam-
Biophys Res Commun. 2002;294:287–292.
matory cytokine activity after menopause. Endocr Rev. 2002;23:
32. Duggan DJ, Bittner M, Chen Y, Meltzer P, Trent JM. Expression
profiling using cDNA microarrays. Nat Genet. 1999;1(suppl):10 –
12. The Eye Disease Case-Control Study Group. Risk factors for neo-
vascular age-related macular degeneration. Arch Ophthalmol.
33. Sharon D, Blackshaw S, Cepko CL, Dryja TP. Profile of the genes
expressed in the human peripheral retina, macula, and retinal
13. Smith W, Mitchell P, Wang JJ. Gender, oestrogen, hormone re-
pigment epithelium determined through serial analysis of gene
placement and age-related macular degeneration: results from the
expression (SAGE). Proc Natl Acad Sci USA. 2002;99:315–320.
Blue Mountains Eye Study. Aust NZ J Ophthalmol. 1997;25(suppl
34. Matsumoto T, Tsurumoto T. Serum YKL-40 levels in rheumatoid
arthritis: correlations between clinical and laboratory parameters.
14. Smith W, Assink J, Klein R, Mitchell P, et al. Risk factors for
Clin Exp Rheumatol. 2001;19:655– 660.
age-related macular degeneration: pooled findings from three con-
35. Johansen JS, Christoffersen P, Moller S, et al. Serum YKL-40 is
tinents. Ophthalmology. 2001;108:697–704.
increased in patients with hepatic fibrosis. J Hepatol. 2000;32:
15. Nonaka A, Kiryu J, Tsujikawa A, et al. Administration of 17beta-
estradiol attenuates retinal ischemia-reperfusion injury in rats. In-
36. Cintin C, Johansen JS, Christensen IJ, Price PA, Sorensen S, Nielsen
vest Ophthalmol Vis Sci. 2000;41:2689 –2696.
HJ. Serum YKL-40 and colorectal cancer. Br J Cancer. 1999;79:
16. Suzuma I, Mandai M, Takagi H, et al. 17--Estradiol increases VEGF
receptor-2 and promotes DNA synthesis in retinal microvascularendothelial cells. Invest Ophthalmol Vis Sci. 1999;40:2122–2129.
37. Johansen JS, Baslund B, Garbarsch C, et al. YKL-40 in giant cells
and macrophages from patients with giant cell arteritis. Arthritis
17. Miyamoto N, Mandai M, Takagi H,et al. Contrasting effect of estro-
gen on VEGF induction under different oxygen status and its role
in murine ROP. Invest Ophthalmol Vis Sci. 2002;43:2007–2014.
38. Boot RG, van Achterberg TA, van Aken BE, et al. Strong induction
18. Hakala BE, White C, Recklies AD. Human cartilage gp-39, a major
of members of the chitinase family of proteins in atherosclerosis:
secretory product of articular chondrocytes and synovial cells, is a
chitotriosidase and human cartilage gp-39 expressed in lesion
mammalian member of a chitinase protein family. J Biol Chem.
macrophages. Arterioscler Thromb Vasc Biol. 1999;19:687– 694.
39. Johansen JS, Olee T, Price PA, Hashimoto S, Ochs RL, Lotz M.
19. Rehli M, Krause SW, Andreesen R. Molecular characterization of
Regulation of YKL-40 production by human articular chondro-
the gene for human cartilage gp-39 (CHI3L1), a member of the
cytes. Arthritis Rheum. 2001;44:826 – 837.
chitinase protein family and marker for late stages of macrophage
40. Rakoczy PE, Lai CM, Baines M, Di Grandi S, Fitton JH, Constable IJ.
differentiation. Genomics. 1997;43:221–225.
Modulation of cathepsin D activity in retinal pigment epithelial
20. De Ceuninck F, Gaufillier S, Bonnaud A, Sabatini M, Lesur C,
cells. Biochem J. 1997;324:935–940.
Pastoureau P. YKL-40 (cartilage gp-39) induces proliferative events
41. Rochefort H, Chalbos D, Cunat S, Lucas A, Platet N, Garcia M.
in cultured chondrocytes and synoviocytes and increases glycos-
Estrogen regulated proteases and antiproteases in ovarian and
aminoglycan synthesis in chondrocytes. Biochem Biophys Res
breast cancer cells. J Steroid Biochem Mol Biol. 2001;76:119 –124.
42. Zhang D, Lai MC, Constable IJ, Rakoczy PE. A model for a blinding
21. Malinda KM, Ponce L, Kleinman HK, Shackelton LM, Millis AJ.
eye disease of the aged. Biogerontology. 2002;3:61– 66.
Gp38K, a protein synthesized by vascular smooth muscle cells,
43. Miralles F, Gaudelet C, Cavailles V, Rochefort H, Augereau P.
stimulates directional migration of human umbilical vein endothe-lial cells. Exp Cell Res. 1999;250:168 –173.
Insensitivity of cathepsin D gene to estradiol in endometrial cells isdetermined by the sequence of its estrogen responsive element.
22. Verheijden GF, Rijnders AW, Bos E, et al. Human cartilage glyco-
protein-39 as a candidate autoantigen in rheumatoid arthritis. Ar-Biochem Biophys Res Commun. 1994;203:711–718. thritis Rheum. 1997;40:1115–1125.
44. Sobel MI, Winkel CA, Macy LB, Liao P, Bjornsson TD. The regula-
23. Vos K, Steenbakkers P, Miltenburg AM, et al. Raised human carti-
tion of plasminogen activators and plasminogen activator inhibitor
lage glycoprotein-39 plasma levels in patients with rheumatoid
type 1 in endothelial cells by sex hormones. Am J Obstet Gynecol.
arthritis and other inflammatory conditions. Ann Rheum Dis.
45. Srivasta RAK, Srivasta N, Averna M, et al. Estrogen up-regulates
24. Johansen JS, Stoltenberg M, Hansen M, et al. Serum YKL-40 con-
apolipoprotein E (ApoE) gene expression by increasing ApoE
centrations in patients with rheumatoid arthritis: relation to dis-
mRNA in the translating pool via the estrogen receptor ␣-mediated
ease activity. Rheumatology. 1999;38:618 – 626.
pathway. J Biol Chem. 1997;272:33360 –33366.
Diclofena Pain/in ammation in RA, acute gout, post-op, fever/pain in children, dysmenorrhoea, migraine. 1. 2-1 ,8g daily inSEs. GI discomfort/bleeding (should take with meals), hypersensitivity, renal failureFrom the BNF. Do not rely on this information for prescribing. CI. Hypersensitivity to any NSAID, severe heart failure. COX2s are CI in some CVS dis-eases now. NSAIDs may interfere with r
CONTRALORÍA MUNICIPAL DE VALLEDUPAR “De la mano contigo vigilamos los bienes de todos” DESCPACHO CONTRALORA Nit: 892.300.310-2 INFORME FINAL DE AUDITORÍA GUBERNAMENTAL CON ENFOQUE INTEGRAL MODALIDAD ESPECIAL HOSPITAL EDUARDO ARREDONDO DAZA ESTADOS CONTABLES VIGENCIA 2010. CONTRALORIA MUNICIPAL DE VALLEDUPAR Mayo de 2011 ¡De la
 Effect of Estrogens on Retinal YKL-40
Effect of Estrogens on Retinal YKL-40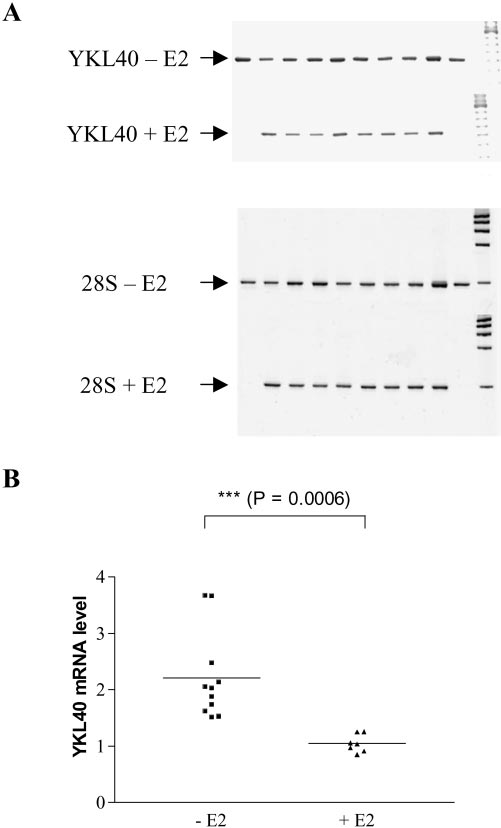 Rakic et al.
Rakic et al.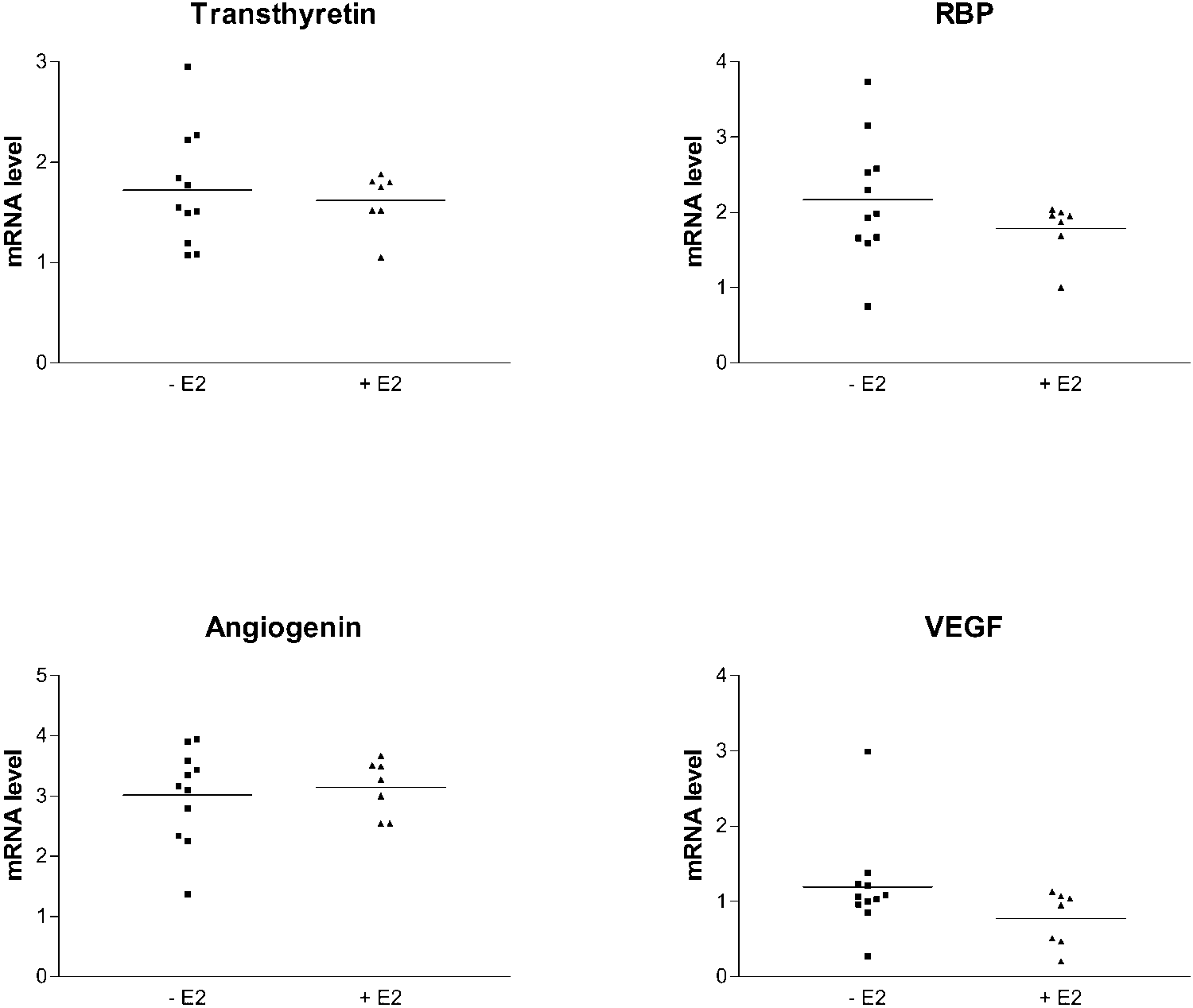 Effect of Estrogens on Retinal YKL-40
Effect of Estrogens on Retinal YKL-40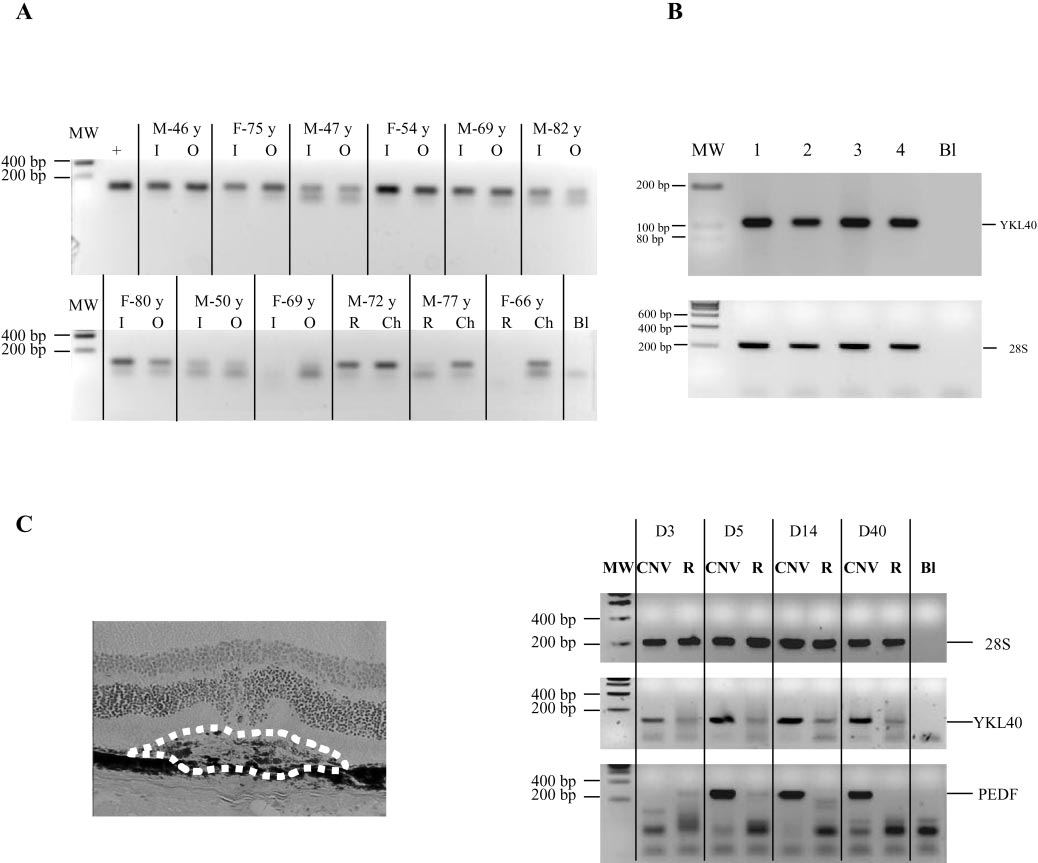 Effect of Estrogens on Retinal YKL-40
Effect of Estrogens on Retinal YKL-40