Levitra has a minimal amount of contraindications which has increased its popularity wellbutrin xl uk You can buy quality certified medications from us at an affordable price.
The coexistence
What are social insects telling us about aging?
Abstract
Research on aging in social insects has progressed much more than has been generally acknowledged. Here I review what I think are the four greatest contributions of social insect work to the field of aging research with the hope of high-lighting the truly exciting discoveries being made. These include the reversal of the fecundity / lifespan and size / lifespan trade-offs due to the evolution of sociality, that social environment can reverse the effects of aging, the contribution of social insect work to the overturning of the free radical theory of aging, and the discovery of vitellogenin as an im-portant protein for longevity. All of these discoveries have important ramifications for human and mammalian aging.
Key words: Aging, ant, Formicidae, bee, free radical theory, longevity, social insects, vitellogenin, review.
Myrmecol. News 13: 103-110 (online xxx 2008) ISSN 1994-4136 (print), ISSN 1997-3500 (online)
Received 13 November 2009; revision received 9 December 2009; accepted 10 December 2009
Dr. Joel D. Parker, School of Biological Sciences, University of Southampton, Southampton, SO16 7PX, UK. E-mail: [email protected]Introduction
Whenever I tell colleagues or others how the queen of one
impact of social insects on biomedical aging research. The
of the most common ants here in Britain (Lasius niger)
four most biomedically relevant results for aging from re-
can live nearly 30 years (KUTTER & STUMPER 1969), the re-
search in social insects are: 1) social evolution can reverse
sponse is always the same; they are amazed. Those who
the assumed physiological trade-offs, 2) the social environ-
have more than a passing interest in aging research imme-
ment dictates and can reverse physiological changes asso-
diately realize that such a small organism with such a long
ciated with aging, 3) social insects' contribution to the over-
lifespan must contain secrets about the evolution of aging.
throw of the free radical theory of aging, and 4) the multi-
Clearly, organisms such as eusocial queens must have some
way of reversing the usual fecundity / lifespan and size /
Social evolution is associated with the reversal of
lifespan trade-offs (see Box 1). Moreover, this reversal and
fecundity / lifespan and size / lifespan trade-offs
extreme longevity correlates with the evolution of eusoci-ality in termites, bees and ants (KELLER & GENOUD 1997).
The primary source for all research on social insect aging
Despite this initial appeal, there are many impediments
is the phylogenetic study of KELLER & GENOUD (1997).
to the study of aging in social insects. First and foremost is
The authors used a phylogenetic analysis to test the hypo-
that aging research is directed at human health and wel-
thesis that the evolution of eusociality correlates with evo-
fare. This has created a hierarchy in funding and research
lution of a long lifespan. The results were as statistically
focus with humans and translational research on top, fol-
significant as they are visually impressive (Fig. 1). That
lowed by mouse, rat and other mammals as the main re-
sociality can increase lifespan can be argued for other so-
search systems, and the invertebrate models of Drosophila
cial animals including humans (CAREY 2001). In general, so-
melanogaster and Caenorhabditis elegans of academic in-
ciality based on kin selection predicts a tendency to evolve
terest. Compared to the collective power of these model
longer lifespans (BOURKE 2007). In most cases of evolved
systems, long lived and challenging non-model systems like
increased lifespan, the evolutionary causes involved low-
ants and bees might seem destined always to be several steps
ered extrinsic mortality for the reproductives in the social
behind. However, the reality is that the results coming out
group. Social groups tend to protect the reproductives from
of social insect work on aging are not just keeping up with
predation while maintaining a more benign environment,
the aging research field, but taking the lead as well. The
thus reducing mortality from predators and stress.
challenge for those of us working in social insects is to get
Aging patterns and social structure are intimately con-
the biomedical research enterprise to notice our most trans-
nected in social organisms with social structure frequently
determining the evolution of aging trajectories. This has
There have been many comprehensive reviews of aging
been modeled and demonstrated for worker age polyethism
in social insects over the last several years (RUEPPELL &
in social insects (BESHERS & al. 2001, TOFILSKI 2002,
al. 2004, JEMIELITY & al. 2005, KELLER & JEMIELITY 2006,
TRIPET & NONACS 2004, TSUJI & TSUJI 2005). However,
BOURKE 2007, REMOLINA & al. 2007, HEINZE & SCHREMPF
the evolution of extreme long life observed in ants and bees
2008, MUNCH & al. 2008). Instead of just recovering this
occurs primarily with female reproductives. In ants this cre-
area, I wish to take a practical perspective and highlight the
ates an extreme difference in lifespan between queens (de-
cades), workers (years) and males (months) (SEELEY 1978,
HÖLLDOBLER & WILSON 1990, HARTMANN & HEINZE
2003). Sometimes overlooked is the exceptional long life
Damage accumulation: the observed accumulation of
of workers (up to several years) (GODZINSKA & al. 1999)
molecular and physical damage to an organism with
which is short compared to queens but is extremely long
lived for their size. Interestingly, workers would seem to
Extrinsic mortality: mortality caused by predation, di-
be under the same evolutionary pressures to evolve long life
sease, starvation, accidents or other environmental causes
as queens, but only until they start to forage. This indi-cates that extrinsic mortality is once again the overriding
Free radicaltheory of aging: the hypothesis that reac-
tive oxygen species cause molecular and cellular damage
The reversals of trade-offs also extend to sexual con-
flict but for different reasons. Mating in insects frequently
Immunity / longevity trade-off: the hypothesis that ro-
reduces female lifespan (CHAPMAN & al. 1995, PROMISLOW
bustness of the immune response trades off against life-
2003), but the reverse was found in the ant Cardiocondyla
span. One can only be increased at a cost to the other.
obscurior where mating increases queen lifespan (SCHREMPF
Intrinsic aging: increase with time in fragility or pro-
& al. 2005). Kin selection and the structure of eusocial col-
bability of dying due to internal causes
onies can favor workers to replace the queen when the
Intrinsic mortality: mortality caused by organ failure,
queen's fertility begins to decline (LEE 2003, BOURKE
cancer or other causes internal to the animal's body
2007). This can happen when a queen's fecundity falls enough that the worker will pass on more of her own
Fecundity / lifespan trade-off: the hypothesis that life-
genes to the next generation by replacing the queen. In-
span and reproductive effort must trade-off. Increasing
deed, through kin selection, the queen herself may favor
one requires decreasing the other. Also called the long-
colony turn-over to a related sister or daughter after the
queen's own fecundity has fallen to some level. The timing
Programmed aging: the idea that organisms are gene-
may also depend on how resources such as rank and terri-
tically determined to grow old and die. Sometimes used
tory may be inherited among kin. There may be a conflict
in the timing of the colony turn-over if the worker's favored
Rate-of-living theory of aging: the hypothesis that an
take-over time comes before the queen's favored hand-over
organism's metabolic rate, sometimes scaled for size, de-
time (BOURKE 2007). These sorts of intergenerational trans-
termines the rate of aging. In the past, this was thought
fer situations provide a rich set of conditions and examples
to happen through controlling the rate of free radical pro-
for studying senescence under varying social and kin selec-
The observed reversal of the fecundity / lifespan trade-
Size / lifespan trade-off: the hypothesis that size and
off seen in many reproductive social insects is an unex-
lifespan trade-off. The original idea is that physics de-
pected and completely revolutionary finding from a mo-
termines the relationship between size and metabolism
lecular and physiological point of view. Until such re-
and larger animals with lower metabolic rate experience
versals were documented and studied, the trade-offs were
assumed more or less universal physiological constraints
on longevity because they were observed across all of the
regulator of cell metabolism controlling many aspects of
short-lived model systems. Exceptions were noticed, but
protein synthesis. The small workers and large queens were
now they can be theoretically explained and it can be
found to be associated with lower and higher TOR activity,
proven that many of these trade-offs are physiologically
respectively. This is the reverse of the expected TOR ac-
malleable to ecological and evolutionary forces.
tivities based on the lifespans (STANFEL & al. 2009). The
These evolved intrinsic aging differences among castes
free radical theory predicts that higher levels of TOR acti-
offer an opportunity to test biochemical hypotheses and re-
vity would lower lifespan by creating more metabolic ac-
sults from the short-lived model systems. The results have
tivity and more free radical damage accumulation. Thus
been surprising, sometimes supporting and sometimes con-
taken together, the biochemical nature of the difference
tradicting the idea that aging will be correlated with the
in intrinsic aging among castes is raising doubts about the
accumulated damage from free radicals. Comparing Super-
free radical theory just as model systems are.
oxide Dismutase (SOD) across the sexes and castes found
One under-exploited aspect of caste-specific lifespans
that the longest-lived castes did not have the most anti-
is the differences among polymorphic sterile worker castes
oxidant protection contradicting the free radical theory of
in ants. CHAPUISAT & KELLER (2002) showed that large
aging (PARKER & al. 2004a). This work also led to the dis-
workers were shorter-lived than small workers in a weaver
covery that insects possessed the extra-cellular form of
ant and argued that it was consistent with extrinsic mor-
SOD (PARKER & al. 2004b). Other molecular work com-
tality acting on the sterile individuals. In this case, there is
paring queens and workers showed the same trend with
reversal of the size / lifespan correlation for sterile wor-
antioxidants in honey bees (CORONA & al. 2005, CORONA
kers. This hypothesis would depend upon some connection
& ROBINSON 2006). Characterizing telomeres across sex
between the mortality rates of the large and small castes
and castes in the ant Lasius niger found both supportive
and the mortality rate of the colony which is the level se-
and contradictory results for the free radical theory (JEMI-
lection must be acting. The relationship of these various lev-
ELITY & al. 2007). More fundamental differences in caste
els acting across colony and different types of sterile wor-
metabolism were found with the TOR pathway (PAGE &
kers is one place where programmed aging could occur.
AMDAM 2007, PATEL & al. 2007) which is a fundamental
is potential for future work to examine the biochem-
al. 2006, WOLSCHIN & AMDAM 2007a, b). Although con-firming fundamental differences between the subcastes in many basic pathways, these studies demonstrate the clas-sic limitation with proteomic and genomic aging studies. The typical experiment is to compare a long-lived and short-lived organism and the result is that the same basic meta-bolic pathways are usually affected. These include the in-sulin, TOR, and stress pathways. These are the three main biochemical processes that appear in many studies on aging
(BROUGHTON & PARTRIDGE 2009, GREWAL 2009, PAPA-CONSTANTINOU 2009, STANFEL & al. 2009), but also so
Fig. 1: The relative reproductive lifespan of various in-
fundamental to regulating metabolism that any major change
sect taxa showing the effect of eusociality. Details on the
in the organism's metabolism will likely perturb them. The
order Hymenoptera are given for several families because
important result is that the subcastes are different at a
eusociality evolved independently in the genus Apis and
basic metabolic level. This is a key lesson for anyone doing
in Formicidae (ants). Reprinted by permission from Mac-
an "omics" experiment on aging. Thus far the results have
millan Publishers Ltd: Nature (KELLER & GENOUD 1997),
been too general to be interpreted by themselves and have
proven most valuable as preliminary work for future re-
ductionistic studies (WHITFIELD & al. 2006, GRAFF & al.
istry of the size / lifespan trade-off in such eusocial sys-
2007). Most genomic studies require confirmatory work be-
tems given that the ant with the most genomic resources
fore meaningful results about specific genes and pathways
available (Solenopsis invicta) also has an extreme size poly-
morphism among its sterile workers (WANG & al. 2007,
One particularly relevant result for biomedicine is the
observed restoration of an age related decline in the im-mune system (AMDAM & al. 2004, 2005). It is well estab-
Social environment can change intrinsic
lished that immune responses change over life time and are
aging patterns in social insects
dependent on the behaviors (foraging or inside workers)
Intrinsic aging rates can be dramatically altered by social
of the worker bees and ants (DOUMS & al. 2002, BOCHER
environment. Honey bees in particular have provided many
& al. 2007, MORET & SCHMID-HEMPEL 2009). Immune
insights into the physiology of the plastic responses of in-
function declines with age in bumble bees (DOUMS & al.
trinsic aging to social environment. For example, the size
2002) but stays the same or even increases in an ant (BO-
of the colony affects individual aging rates (RUEPPELL &
CHER & al. 2007). In honey bees, it drops when bees begin
al. 2009). In the trade-off between colony and individual,
to forage but can be restored when these bees are switched
it is the individual situation that determines how the bees
back to hive tasks (AMDAM & al. 2005). Thus, the decline
age (RUEPPELL & al. 2008). Environment has been shown
in immune function with age is reversible and not the re-
to be more important in determining how honey bees age
sult of permanent damage accumulation. Indeed, the pros-
than the actual time the bee has been alive (RUEPPELL &
pect of rejuvenating the human immune system is one seri-
al. 2007b). The key seems to be when workers make the
ous idea being considered to combat human aging (DORSH-
transition to foraging and their mortality rates rise dramati-
cally (RUEPPELL & al. 2007a, WOYCIECHOWSKI & MORON
The idea that social interactions are important in aging
has recently been tested in a Drosophila system which
In honeybees, workers that develop in the autumn are
produced a genuinely baffling result (RUAN & WU 2008).
long-lived whereas workers developing in the spring are
The authors cited the above honey bee work as the justifi-
short-lived (MAURIZIO 1950, BECERRA-GUZMAN & al.
cation and inspiration for these experiments. Short-lived fruit
2005). These subcastes can be determined by diet during
flies with a cytoplasmic Cu-Zn SOD mutation were placed
development and can live up to a year. These observations
in an environment with long-lived flies and with younger
combined with the ability to revert foragers back to brood
short-lived flies. Living with younger flies caused the mu-
care by altering their environment (ROBINSON & al. 1992)
tant lines to recover a significant portion of the wild type
provide manipulative tests for examining the changes as-
fly lifespan compared to controls. The authors then went
sociated with aging. This ability to revert aging pheno-
through a series of carefully controlled experiments (con-
types in this system has shown vitellogenin, Juvenile Hor-
trolling food, decapitation, clipping wings, making the
mone, and the insulin pathways as the regulators control-
SOD mutants deaf, co-housing in the dark) to show that
ling and responding to the two subcastes (AMDAM & OM-
the effect was mediated by behavior. How this works is yet
HOLT 2003, AMDAM & al. 2004, GUIDUGLI & al. 2005,
to be explored, but the conclusion is that social environ-
AMDAM & al. 2007, CORONA & al. 2007, NELSON & al.
ment effects individual aging rates, even in what are usu-
2007, MUNCH & al. 2008). The link to the insulin path-
way is particularly intriguing as it is one of the main path-
This idea that organisms age depending on their social
ways associated with aging in model systems. Even im-
interactions has the potential to be one of the most clini-
mune system function and the ability to learn could be re-
cally relevant transferable findings to human aging. Hu-
versed in these types of experiments (AMDAM & al. 2004,
mans are social animals and almost certainly under similar
AMDAM & PAGE 2005, BEHRENDS & al. 2007).
selective pressures for increased longevity as social insects.
Proteomic studies have been done with the winter/sum-
Indeed social environment has been shown to have an ef-
mer bee system comparing the subcastes (SCHIPPERS &
fect on aging in humans although the effect is mediated by
the type of interactions (ROHR & LANG 2009). Clinical re-
ant nests as predicted by the old free radical theory. The
search has tended to separate positive and negative social
evidence does point in both directions, even in single stud-
interactions with subsequent positive and negative effects
ies as seen in ant telomeres where males have shorter te-
on lifespan. While there are ethical barriers to the most ob-
lomeres than workers and queens as predicted by the old
vious replication of the honey bee results in humans, we
free radical theory, but workers and queens have telomeres
can test whether persons with child-care responsibilities
of about the same length contradicting the theory (JEMIE-
later in life are aging differently (mimicking the shift from
LITY & al. 2007). The simple free radical theory and dam-
forager to nurse bee). Early human studies suggest mixed
age accumulation can not explain the contradicting cases
results depending on various external stressors (BURNETTE
2000). It may be that honey bees are an ideal case where
The idea that aging is not caused by accumulated damage
there are no negative effects on lifespan by virtue of evo-
can be very difficult to accept. There is a common every-
lutionary pressures on the sterile castes. Coming from the
day example that explains how an organism can age without
human clinical direction, the effect of quality of the inter-
accumulating physical damage. Consider how many
actions in humans might be tested in less socially evolved
computer failures you have had in your lifetime. By failure,
insects like wasps. How does the rate of aging change with
consider any time your machine locked up (needing a re-
respect to position in a dominance hierarchy (TSUJI & TSUJI
start) which would be the equivalent of death in a biolo-
2005), amount of violent interactions, and just as important-
gical system. Now consider how many have been due to
ly, what molecular mechanisms might underpin any change?
hardware failures and how many due to software problems. In almost all cases computers are far more likely to lose
Social insects and the death of the
function due to their internal state than due to the physical
free radicaltheory of aging
breakdown of a component. Thus, computers and other
The old free radical theory, that accumulation of oxidative
complex systems like living organisms can fail globally be-
damage causes aging (HARMAN 1956), is in the midst of be-
cause they arrive in an inappropriate state, without any spe-
ing overthrown. There is now overwhelming evidence that
cific subsystem failure (all the parts can be functional, but
aging is not caused by a simple accumulation of free rad-
the whole is not). Such bad states in organisms and cells
ical damage on macromolecules (BLAGOSKLONNY 2008,
would involve changes to fundamental metabolic states or
BRINK & al. 2009, GEMS & DOONAN 2009, PEREZ & al.
developmental programs and might be expected to cause
2009). Instead, free radicals appear to be signaling mole-
stress and damage leading to the observed correlations of
cules related to stress as well as participants in fundamen-
tal metabolic processes (BLAGOSKLONNY 2008, JUAREZ &
How then can an organism age when the cause is not
al. 2008, MAGLIARO & SALDANHA 2009). Part of this is
accumulated damage? There are at least three recently pro-
the regulation of damaged molecule turn-over (CHEN & al.
posed and one older relatively unknown theory for how or-
2009). Hence, the accumulation of damage and the increased
ganisms can age without accumulating damage being the
mortality rate that defines aging appear to be caused by
cause. Demetrius (DEMETRIUS 2004, BRINK & al. 2009)
fundamental changes in regulatory and developmental path-
proposes a metabolic instability hypothesis that the rate of
ways (BUDOVSKAYA & al. 2008). These in turn are likely
living coupled with demographic factors leads to metabolic
effecting the accumulation of oxidative damage, autophagy
instability causing aging. BLAGOSKLONNY (2008) describes
and cellular repair mechanisms. What is happening to the
what he calls "quasi-programmed aging" based on TOR-re-
free radical theory of aging is a reversal of cause and ef-
lated pathways being switched on by mistake over time.
fect with damage looking more like the effect of aging pro-
BUDOVSKAYA & al. (2008) suggest that aging is caused by
cesses as opposed to the cause of aging (BLAGOSKLONNY
developmental programs becoming inappropriately acti-
vated with age and labels it the developmental drift theory
Much support for the paradigm shift is coming from
of aging. In 1978, Robert Rosen proposed the feedforward
studies in social insects. The already stated reversal of the
theory of aging arguing that all complex systems with feed-
fecundity / lifespan trade-off in queens supports the new
forward pathways (predictive models, not self-correcting)
interpretation. Levels of the antioxidant Superoxide Dis-
will inevitably fail at the global level without the need for
mutase were higher in short-lived ant castes contrary to pre-
any specific subsystem failure (ROSEN 1978). Basically,
dictions of the old free radical theory (PARKER & al. 2004b)
his idea is that every complex system will inevitably get
while much more extensive genomic work found the same
into a state incompatible with function as in the computer
trends in honey bee (CORONA & al. 2005, CORONA & RO-
example above. Although these theories have not been fully
BINSON 2006). Additionally, free radical damage was found
developed nor synthesized, they all share the characteris-
not to be associated with age in honey bee (SEEHUUS &
tics of being systems level explanations and can explain the
al. 2006a) despite the antioxidant vitellogenin (SEEHUUS &
results that the free radical theory and damage accumula-
al. 2006b). Heatshock-gene expression does not agree with
the simple free radical theory as some important heatshock-
The important point is that the experimental evidence
gene expression levels were observed to decline with age
currently supports the notion that higher system level ef-
in worker honey bees (AAMODT 2008). Finally, brain de-
fects are more important drivers of aging than physical dam-
velopment and complexity increase with age in both bees
age accumulation. Social insect aging results are support-
(FARRIS & al. 2001) and ants (SEID & al. 2005, SEID &
ing this emerging paradigm. Social-insect researchers are
al. 2008) while "wear and tear" would predict the opposite.
already used to thinking in terms of emergent properties
There are some correlations in the other direction such as
and systems level phenomena. Hence, we should be in an
the observed lifespan correlation with peroxidation-resis-
intellectually strong position to contribute to this new emerg-
tant lipids (HADDAD & al. 2007) and requeening of wood
The role of vitellogenin in aging
mainstream biomedical aging-research community. As I see
it, the key points of entry are 1) the evolution of reversals
The significance of the discovery of vitellogenin as a major
of physiological constraints on lifespan, 2) socially rever-
player in aging (AMDAM & OMHOLT 2003, OMHOLT & AM-
sible aging phenotypes, 3) the paradigm shift in the free
DAM 2004, NELSON & al. 2007, AMDAM & al. 2009) has
radical theory, and 4) the role of vitellogenin in aging. We
yet to be recognized for its true potential significance. This
have the advantage of natural and easily manipulated sys-
is an excellent example of how a discovery in social in-
tems that by definition far outstrip the capabilities of any
sects is slow to penetrate into the more mainstream aging
model system to explore these areas. Indeed the weaknes-
field. Vitellogenin is thought to be related to mammalian
ses of our social experimental systems can be overcome
low density lipoproteins (LDL) (SAPPINGTON & RAIKHEL
by testing in mainstream model systems through closer
1998, SMOLENAARS & al. 2007). LDL reactions in the vas-
collaboration with biomedical researchers. Only when these
cular endothelium of humans involve oxidative stress, in-
results are replicated in established laboratory mammalian
flammation, and programmed cell death and are thought to
models will our results finally be transferable to human
be major players in human cardiovascular disease (SIMA &
al. 2009, WILENSKY & MACPHEE 2009). Understanding the
Social behavior is the ultimate determinate of long-
dynamics of vitellogenin might yield insights into human
evity in humans, and social insects offer the only real op-
tion for understanding the evolutionary forces underpinning
Social insects appear unique in that the adult workers
sociality and lifespan. Making this connection, and always
can be expressing high amounts of vitellogenin in a com-
looking towards the general principles at both the proxi-
plex regulatory network with juvenile hormone (JH) (GUI-
mate and ultimate level can not help but raise the profile of
DUGLI & al. 2005, AMDAM & al. 2007). In sterile workers,
JH can suppress vitellogenin and vitellogenin in turn can
Acknowledgements
suppress JH in what is called the double repressor hypothe-sis (AMDAM & OMHOLT 2003, OMHOLT & AMDAM 2004).
I thank Karen Parker for reading several versions of this
The switch-over from nurse bee (high vitellogenin) to forag-
manuscript. I also thank the patient editors at Myrmecolo-
er (low vitellogenin) is then associated with a shift to more
gical News and two anonymous reviewers whose comments
rapid aging (FLURI & al. 1981, 1982).
Vitellogenin acts as an antioxidant in honey bees by be-
Zusammenfassung
coming irreversibly carbonylated (SEEHUUS & al. 2006b). Vitellogenin scavenges oxidants by essentially offering it-
Die Erforschung der Alterung bei sozialen Insekten hat
self as a noncatalytic target for the damage. On the sur-
viel größere Fortschritte gemacht als landläufig bekannt.
face, this seemed a perfect prediction for the damage accu-
Hier biete ich einen Überblick über die vier meiner Ansicht
mulation version of the free radical theory, but the meas-
nach größten Beiträge der Forschung an sozialen Insekten
ured accumulation of damage was better explained by so-
zum Gebiet der Alterungsforschung, in der Hoffnung, die
wahrlich fesselnden Entdeckungen, die gemacht wurden,
The most novel finding is that vitellogenin is a zinc car-
herauszustellen. Dies sind die Umkehr der Trade-offs von
rier and this function might be associated with the immu-
Fruchtbarkeit und Lebensspanne und von Größe und Le-
nity / longevity trade-off (AMDAM & al. 2004). The signi-
bensspanne in Folge der Evolution von Sozialität, die Er-
ficance of this discovery urgently needs further exploration,
kenntnis, dass die soziale Umwelt die Effekte des Alterns
especially as it does not appear that zinc is associated with
umkehren kann, der Beitrag der Forschung an sozialen In-
the antioxidant effect of vitellogenin. The highly expressed
sekten zum Kippen der Freie-Radikale-Theorie des Alte-
zinc containing enzymes Cu-Zn SODs are major enzymes
rungsprozesses sowie die Entdeckung von Vitellogenin als
implicated in aging. Also, a well known age-related neuro-
ein für die Langlebigkeit wichtiges Protein. Alle diese Ent-
degenerative disease, familial amyotrophic lateral sclerosis
deckungen sind ausgesprochen relevant in Hinblick auf das
(or motor neuron disease) and other neurological damage
Altern beim Menschen und bei anderen Säugetieren.
can be caused by changes in Cu-Zn SOD that destabilize the zinc-binding ability of the enzyme (E
References
ROBERTS & al. 2007, SMITH & LEE 2007, DING & DOK-
AGOSTI, D. 1990: Review and reclassification of Cataglyphis
HOLYAN 2008). Furthermore, zinc is required in important
(Hymenoptera, Formicidae). – Journal of Natural History 24:
gene regulatory pathways (zinc fingers) and interacts with
the NMDA receptor which is important for learning (KLUG
AAMODT, R.M. 2008: The caste- and age-specific expression sig-
2005, KANDEL 2009, PAOLETTI & al. 2009). The vitello-
nature of honeybee heat shock genes shows an alternative splic-
genin results are pointing to the hypothesis that the move-
ing-dependent regulation of hsp90. – Mechanisms of Ageing
ment and correct localization of zinc may be important to
AMDAM, G.V., AASE, A., SEEHUUS, S.C., FONDRK, M.K., NOR-
Thus, vitellogenin is one of the most exciting molecular
BERG, K. & HARTFELDER, K. 2005: Social reversal of immuno-senescence in honey bee workers. – Experimental Gerontol-
leads for mainstream model system research in aging com-
ing from social insects today. I predict that we will see this
AMDAM, G.V., NILSEN, K.A., NORBERG, K., FONDRK, M.K. &
discovery being explored in flies and mammalian systems
HARTFELDER, K. 2007: Variation in endocrine signaling under-
lies variation in social life history. – American Naturalist 170:
The future
AMDAM, G.V. & OMHOLT, S.W. 2003: The hive bee to forager
One of the main goals of those working in aging research
transition in honeybee colonies: the double repressor hypo-
in social insects has to be transferring our results to the
thesis. – Journal of Theoretical Biology 223: 461-464.
AMDAM, G.V. & PAGE, R.E., Jr. 2005: Intergenerational trans-
CORONA, M. & ROBINSON, G.E. 2006: Genes of the antioxidant
fers may have decoupled physiological and chronological age
system of the honey bee: Annotation and phylogeny. – Insect
in a eusocial insect. – Ageing Research Reviews 4: 398-408.
AMDAM, G.V., RUEPPELL, O., FONDRK, M.K., PAGE, R.E. & NEL-
CORONA, M., VELARDE, R.A., REMOLINA, S., MORAN-LAUTER,
SON, C.M. 2009: The nurse's load: Early-life exposure to brood-
A., WANG, Y., HUGHES, K.A. & ROBINSON, G.E. 2007: Vitello-
rearing affects behavior and lifespan in honey bees (Apis mel-
genin, juvenile hormone, insulin signaling, and queen honey
lifera). – Experimental Gerontology 44: 467-471.
bee longevity. – Proceedings of the National Academy of Sci-
AMDAM, G.V., SIMOES, Z.L.P., HAGEN, A., NORBERG, K., SCHRO-
ences of the United States of America 104: 7128-7133.
DER, K., MIKKELSEN, O., KIRKWOOD, T.B.L. & OMHOLT, S.W.
DEMETRIUS, L. 2004: Caloric restriction, metabolic rate, and en-
2004: Hormonal control of the yolk precursor vitellogenin re-
tropy. – Journals of Gerontology Series A: Biological Sciences
gulates immune function and longevity in honeybees. – Ex-
DING, F. & DOKHOLYAN, N.V. 2008: Dynamical roles of metal
BECERRA-GUZMAN, F., GUZMAN-NOVOA, E., CORREA-BENITEZ, A.
ions and the disulfide bond in Cu, Zn superoxide dismutase
& ZOZAYA-RUBIO, A. 2005: Length of life, age at first forag-
folding and aggregation. – Proceedings of the National Academy
ing and foraging life of Africanized and European honey bee
of Sciences of the United States of America 105: 19696-19701.
(Apis mellifera) workers, during conditions of resource abund-
ance. – Journal of Apicultural Research 44: 151-156.
ORSHKIND, K., MONTECINO-RODRIGUEZ, E. & SIGNER, R.A.J.
2009: The ageing immune system: Is it ever too old to become
BEHRENDS, A., SCHEINER, R., BAKER, N. & AMDAM, G.V. 2007:
young again? – Nature Reviews Immunology 9: 57-62.
Cognitive aging is linked to social role in honey bees (Apis mellifera). – Experimental Gerontology 42: 1146-1153.
DOUMS, C., MORET, Y., BENELLI, E. & SCHMID-HEMPEL, P. 2002:
Senescence of immune defence in Bombus workers. – Eco-
BESHERS, S.N., HUANG, Z.Y., OONO, Y. & ROBINSON, G.E. 2001:
Social inhibition and the regulation of temporal polyethism in honey bees. – Journal of Theoretical Biology 213: 461-479.
ESTEVEZ, A.G., CROW, J.P., SAMPSON, J.B., REITER, C., ZHUANG,
Y., RICHARDSON, G.J., TARPEY, M.M., BARBEITO, L. & BECK-
BLAGOSKLONNY, M.V. 2008: Aging ROS or TOR. – Cell Cycle 7:
MAN, J.S. 1999: Induction of nitric oxide – dependent apopto-
sis in motor neurons by zinc-deficient superoxide dismutase.
BOCHER, A., TIRARD, C. & DOUMS, C. 2007: Phenotypic plasti-
city of immune defence linked with foraging activity in the
FARRIS, S.M., ROBINSON, G.E. & FAHRBACH, S.E. 2001: Experi-
ant Cataglyphis velox. – Journal of Evolutionary Biology 20:
ence- and age-related outgrowth of intrinsic neurons in the
mushroom bodies of the adult worker honeybee. – Journal of
BOURKE, A.F.G. 2007: Kin selection and the evolutionary theory
of aging. – Annual Review of Ecology, Evolution, and Syste-matics 38: 103-128.
FLURI, P., LÜSCHER, M., WILLE, H. & GERIG, L. 1982: Changes
in weight of the pharyngeal gland and haemolymph titres of
BRINK, T.C., DEMETRIUS, L., LEHRACH, H. & ADJAYE, J. 2009:
juvenile hormone, protein and vitellogenin in worker honey
Age-related transcriptional changes in gene expression in dif-
bees. – Journal of Insect Physiology 28: 61-68.
ferent organs of mice support the metabolic stability theory of aging. – Biogerontology 10: 549-564.
FLURI, P., SABATINI, A.G., VECCHI, M.A. & WILLE, H. 1981: Blood
juvenile-hormone, protein and vitellogenin titers in laying and
BROUGHTON, S. & PARTRIDGE, L. 2009: Insulin/IGF-like signalling,
non-laying queen honeybees. – Journal of Apicultural Research
the central nervous system and aging. – Biochemical Journal
GEMS, D. & DOONAN, R. 2009: Antioxidant defense and aging
BUDOVSKAYA, Y.V., WU, K., SOUTHWORTH, L.K., MIN, J., TE-
in C. elegans: is the oxidative damage theory of aging wrong?
DESCO, P., JOHNSON, T.E. & KIM, S.K. 2008: An elt-3/elt-5/
elt-6 gata transcription circuit guides aging in C. elegans. – Cell 134: 291-303.
GODZINSKA, E.J., SZCZUKA, A. & KORCZYNSKA, J. 1999: Maxi-
mum longevity of workers of three ant species under labora-
BURNETTE, D. 2000: Latino grandparents rearing grandchildren
tory conditions (Hymenoptera: Formicidae). – Polskie Pismo
with special needs – effects on depressive symptomatology. –
Journal of Gerontological Social Work 33: 1-16.
GRAFF, J., JEMIELITY, S., PARKER, J.D., PARKER, K.M. & KELLER,
AREY, J.R. 2001: Demographic mechanisms for the evolution
of long life in social insects. – Experimental Gerontoloy 36:
L. 2007: Differential gene expression between long-lived queens
and short-lived workers in the ant Lasius niger. – Molecular Ecology 16: 675-683.
CHAPMAN, T., LIDDLE, L.F., KALB, J.M., WOLFNER, M.F. & PAR-
TRIDGE, L. 1995: Cost of mating in Drosophila melanogaster
REWAL, S.S. 2009: Insulin/TOR signaling in growth and ho-
females is mediated by male accessory gland products. – Na-
meostasis: a view from the fly world. – International Journal
of Biochemistry & Cell Biology 41: 1006-1010.
CHAPUISAT, M. & KELLER, L. 2002: Division of labour influences
GUIDUGLI, K.R., NASCIMENTO, A.M., AMDAM, G.V., BARCHUK,
the rate of ageing in weaver ant workers. – Proceedings of
A.R., OMHOLT, S., SIMOES, Z.L. & HARTFELDER, K. 2005: Vitel-
the Royal Society of London - Series B: Biological Sciences
logenin regulates hormonal dynamics in the worker caste of a
eusocial insect. – Federation of European Biochemical Socie-ties Letters 579: 4961-4965.
CHEN, Y., AZAD, M.B. & GIBSON, S.B. 2009: Superoxide is the
major reactive oxygen species regulating autophagy. – Cell
HADDAD, L.S., KELBERT, L. & HULBERT, A.J. 2007: Extended
Death and Differentiation 16: 1040-1052.
longevity of queen honey bees compared to workers is asso-ciated with peroxidation-resistant membranes. – Experimental
CORONA, M., HUGHES, K.A., WEAVER, D.B. & ROBINSON, G.E.
2005: Gene expression patterns associated with queen honey bee longevity. – Mechanisms of Ageing and Development 126:
HARMAN, D. 1956: Aging: a theory based on free radical and
radiation chemistry. – Journal of Gerontology 11: 298-300.
HARTMANN, A. & HEINZE, J. 2003: Lay eggs, live longer: divi-
PAPACONSTANTINOU, J. 2009: Insulin/IGF-1 and ROS signaling
sion of labor and life span in a clonal ant species. – Evolution
pathway cross-talk in aging and longevity determination. –
Molecular and Cellular Endocrinology 299: 89-100.
HEINZE, J. & SCHREMPF, A. 2008: Aging and reproduction in so-
PARKER, J.D., PARKER, K.M. & KELLER, L. 2004a: Molecular phy-
cial insects – a mini-review. – Gerontology 54: 160-167.
logenetic evidence for an extracellular Cu Zn superoxide dis-
HÖLLDOBLER, B. & WILSON, E.O. 1990: The ants. – The Belknap
mutase gene in insects. – Insect Molecular Biology 13: 587-594.
Press of Harvard University Press, 732 pp.
PARKER, J.D., PARKER, K.M., SOHAL, B.H., SOHAL, R.S. & KEL-
JEMIELITY, S., CHAPUISAT, M., PARKER, J.D. & KELLER, L. 2005:
LER, L. 2004b: Decreased expression of Cu-Zn superoxide dis-
Long live the queen: studying aging in social insects. – Age
mutase 1 in ants with extreme lifespan. – Proceedings of the
National Academy of Sciences of the United States of America
JEMIELITY, S., KIMURA, M., PARKER, K.M., PARKER, J.D., CAO,
X.J., AVIV, A. & KELLER, L. 2007: Short telomeres in short-
PATEL, A., FONDRK, M.K., KAFTANOGLU, O., EMORE, C., HUNT,
lived males: What are the molecular and evolutionary causes?
G., FREDERICK, K. & AMDAM, G.V. 2007: The making of a
queen: TOR pathway is a key player in diphenic caste develop-
JUAREZ, J.C., MANUIA, M., BURNETT, M.E., BETANCOURT, O.,
ment. – Public Library of Science ONE 2: e509.
BOIVIN, B., SHAW, D.E., TONKS, N.K., MAZAR, A.P. & DONATE,
PEREZ, V.I., BOKOV, A., VAN REMMEN, H., MELE, J., RAN, Q.T.,
F. 2008: Superoxide dismutase 1 (sod1) is essential for H2O2-
IKENO, Y. & RICHARDSON, A. 2009: Is the oxidative stress the-
mediated oxidation and inactivation of phosphatases in growth
ory of aging dead? – Biochimica et Biophysica Acta-General
factor signaling. – Proceedings of the National Academy of
Sciences of the United States of America 105: 7147-7152.
PROMISLOW, D. 2003: Mate choice, sexual conflict, and evolution
KANDEL, E.R. 2009: The biology of memory: A forty-year per-
of senescence. – Behavior Genetics 33: 191-201.
spective. – Journal of Neuroscience 29: 12748-12756.
REMOLINA, S.C., HAFEZ, D.M., ROBINSON, G.E. & HUGHES, K.A.
KELLER, L. & GENOUD, M. 1997: Extraordinary lifespans in ants:
2007: Senescence in the worker honey bee Apis mellifera. –
A test of evolutionary theories of ageing. – Nature 389: 958-960.
Journal of Insect Physiology 53: 1027-1033.
KELLER, L. & JEMIELITY, S. 2006: Social insects as a model to
study the molecular basis of ageing. – Experimental Gerontol-
ROBERTS, B.R., TAINER, J.A., GETZOFF, E.D., MALENCIK, D.A.,
ANDERSON, S.R., BOMBEN, V.C., MEYERS, K.R., KARPLUS, P.A. & BECKMAN, J.S. 2007: Structural characterization of zinc-
KLUG, A. 2005: The discovery of zinc fingers and their develop-
deficient human superoxide dismutase and implications for
ment for practical applications in gene regulation. – Proceed-
ALS. – Journal of Molecular Biology 373: 877-890.
ings of the Japan Academy Series B - Physical and Biological Sciences 81: 87-102.
ROBINSON, G.E., PAGE, R.E., STRAMBI, C. & STRAMBI, A. 1992:
Colony integration in honey-bees – mechanisms of behavioral
KUTTER, H. & STUMPER, R. 1969: Hermann Appel, ein leidge-
adelter Entomologe (1892-1966). – Proceedings of the sixth International Congress of the IUSSI (Bern), pp. 275-279.
ROHR, M.K. & LANG, F.R. 2009: Aging well together – a mini-
EE, R.D. 2003: Rethinking the evolutionary theory of aging:
Transfers, not births, shape senescence in social species. – Pro-
ROSEN, R. 1978: Feedforwards and global system failure: a gen-
ceedings of the National Academy of Sciences of the United
eral mechanism for senescence. – Journal of Theoretical Biol-
MAGLIARO, B.C. & SALDANHA, C.J. 2009: Clozapine protects
RUAN, H.Y. & WU, C.F. 2008: Social interaction-mediated life-
PC-12 cells from death due to oxidative stress induced by
span extension of Drosophila Cu/Zn superoxide dismutase
hydrogen peroxide via a cell-type specific mechanism involv-
mutants. – Proceedings of the National Academy of Sciences
ing inhibition of extracellular signal-regulated kinase phos-
of the United States of America 105: 7506-7510.
phorylation. – Brain Research 1283: 14-24.
RUEPPELL, O., AMDAM, G.V., PAGE, R.E., Jr. & CAREY, J.R. 2004:
MAURIZIO, A. 1950: The influence of pollen feeding and brood
From genes to societies. – Science of Aging Knowledge En-
rearing on the length of life and physiological conditions of
RUEPPELL, O., BACHELIER, C., FONDRK, M.K. & PAGE, R.E. 2007a:
MORET, Y. & SCHMID-HEMPEL, P. 2009: Immune responses of
Regulation of life history determines lifespan of worker honey
bumblebee workers as a function of individual and colony age:
bees (Apis mellifera). – Experimental Gerontology 42: 1020-1032.
senescence versus plastic adjustment of the immune function. – Oikos 118: 371-378.
RUEPPELL, O., CHRISTINE, S., MULCRONE, C. & GROVES, L. 2007b:
Aging without functional senescence in honey bee workers. –
MUNCH, D., AMDAM, G.V. & WOLSCHIN, F. 2008: Ageing in a
eusocial insect: molecular and physiological characteristics of life span plasticity in the honey bee. – Functional Ecology 22:
RUEPPELL, O., KAFTANOUGLU, O. & PAGE, R.E. 2009: Honey
bee (Apis mellifera) workers live longer in small than in large colonies. – Experimental Gerontology 44: 447-452.
NELSON, C.M., IHLE, K.E., FONDRK, M.K., PAGE, R.E. & AMDAM,
G.V. 2007: The gene vitellogenin has multiple coordinating
RUEPPELL, O., LINFORD, R., GARDNER, P., COLEMAN, J. & FINE,
effects on social organization. – Public Library of Science
K. 2008: Aging and demographic plasticity in response to
experimental age structures in honeybees (Apis mellifera). –
Behavioral Ecology and Sociobiology 62: 1621-1631.
MHOLT, S.W. & AMDAM, G.V. 2004: Epigenetic regulation of
aging in honeybee workers. – Science of Aging Knowledge
SAPPINGTON, T.W. & RAIKHEL, A.S. 1998: Molecular characte-
ristics of insect vitellogenins and vitellogenin receptors. – In-
PAGE, R.E. & AMDAM, G.V. 2007: The making of a social insect:
sect Biochemistry and Molecular Biology 28: 277-300.
developmental architectures of social design. – Bioessays 29:
SCHIPPERS, M.-P., DUKAS, R., SMITH, R.W., WANG, J., SMOLEN,
K. & MCCLELLAND, G.B. 2006: Lifetime performance in for-
PAOLETTI, P., VERGNANO, A.M., BARBOUR, B. & CASADO, M. 2009:
aging honeybees: behaviour and physiology. – Journal of Ex-
Zinc at glutamatergic synapses. – Neuroscience 158: 126-136.
SCHREMPF, A., HEINZE, J. & CREMER, S. 2005: Sexual coopera-
TRIPET, F. & NONACS, P. 2004: Foraging for work and age-
tion: Mating increases longevity in ant queens. – Current Biol-
based polyethism: the roles of age and previous experience on
task choice in ants. – Ethology 110: 863-877.
SEEHUUS, S.C., KREKLING, T. & AMDAM, G.V. 2006a: Cellular sen-
TSUJI, K. & TSUJI, N. 2005: Why is dominance hierarchy age-
escence in honey bee brain is largely independent of chrono-
related in social insects? The relative longevity hypothesis. –
logical age. – Experimental Gerontology 41: 1117-1125.
Behavioral Ecology and Sociobiology 58: 517-526.
SEEHUUS, S.C., NORBERG, K., GIMSA, U., KREKLING, T. & AMDAM,
WANG, J., JEMIELITY, S., UVA, P., WURM, Y., GRAFF, J. & KELLER,
G.V. 2006b: Reproductive protein protects functionally ste-
L. 2007: An annotated cDNA library and microarray for large-
rile honey bee workers from oxidative stress. – Proceedings
scale gene-expression studies in the ant Solenopsis invicta. –
of the National Academy of Sciences of the United States of
WANG, J., ROSS, K.G. & KELLER, L. 2008: Genome-wide expres-
SEELEY, T.D. 1978: Life history strategy of the honey bee, Apis
sion patterns and the genetic architecture of a fundamental so-
mellifera. – Oecologia 32: 109-118.
cial trait. – Public Library of Science Genetics 4: e1000127.
SEID, M.A., GOODE, K., LI, C. & TRANIELLO, J.F.A. 2008: Age-
WHITFIELD, C.W., BEN-SHAHAR, Y., BRILLET, C., LEONCINI, I.,
and subcaste-related patterns of serotonergic immunoreacti-
CRAUSER, D., LECONTE, Y., RODRIGUEZ-ZAS, S. & ROBINSON,
vity in the optic lobes of the ant Pheidole dentata. – Develop-
G.E. 2006: Genomic dissection of behavioral maturation in
the honey bee. – Proceedings of the National Academy of
SEID, M.A., HARRIS, K.M. & TRANIELLO, J.F.A. 2005: Age-re-
Sciences of the United States of America 103: 16068-16075.
lated changes in the number and structure of synapses in the
WILENSKY, R.L. & MACPHEE, C.H. 2009: Lipoprotein-associated
lip region of the mushroom bodies in the ant Pheidole den-
phospholipase A(2) and atherosclerosis. – Current Opinion in
tata. – Journal of Comparative Neurology 488: 269-277.
SIMA, A.V., STANCU, C.S. & SIMIONESCU, M. 2009: Vascular en-
WOLSCHIN, F. & AMDAM, G.V. 2007a: Comparative proteomics
dothelium in atherosclerosis. – Cell and Tissue Research 335:
reveal characteristics of life-history transitions in a social in-
SMITH, A.P. & LEE, N.M. 2007: Role of zinc in ALS. – Amyo-
WOLSCHIN, F. & AMDAM, G.V. 2007b: Plasticity and robustness
of protein patterns during reversible development in the honey
SMOLENAARS, M.M.W., MADSEN, O., RODENBURG, K.W. & VAN
bee (Apis mellifera). – Analytical and Bioanalytical Chemistry
DER HORST, D.J. 2007: Molecular diversity and evolution of
the large lipid transfer protein superfamily. – Journal of Lipid
WOYCIECHOWSKI, M. & MORON, D. 2009: Life expectancy and
onset of foraging in the honeybee (Apis mellifera). – Insectes
STANFEL, M.N., SHAMIEH, L.S., KAEBERLEIN, M. & KENNEDY,
B.K. 2009: The TOR pathway comes of age. – Biochimica et
WURM, Y., UVA, P., RICCI, F., WANG, J., JEMIELITY, S., ISELI, C.,
Biophysica Acta-General Subjects 1790: 1067-1074.
FALQUET, L. & KELLER, L. 2009: Fourmidable: a database for
TOFILSKI, A. 2002: Influence of age polyethism on longevity of
ant genomics. – BioMed Central Genomics 10: 5.
workers in social insects. – Behavioral Ecology and Sociobiol-ogy 51: 234-237.
11535 Sorrento Valley Blvd, San Diego, CA www.mpexpharma.com Aeroquin (proprietary aerosol formulation of levofloxacin) a reduced risk clinical program; 100% of rights retained• Compelling results from Phase 2b trial in Cystic Fibrosis• Preparing to initiate Phase 3 trial in Cystic Fibrosis Separate broad technology platform focused on efflux pump inhibitors (EPIs) • Coll
Northern International Studies Association Conference “Power, Vision, and Order in World Politics” 23-25 May 2007, University of Southern Denmark, Odense Defining, Allocating, Sharing: Responsibility and Accountability in Global Health Governance Carmen Huckel Research Associate: Graduate Research Programme “Global Challenges” – University of First Draft Only! Pleas
 What are social insects telling us about aging?
Abstract
What are social insects telling us about aging?
Abstract 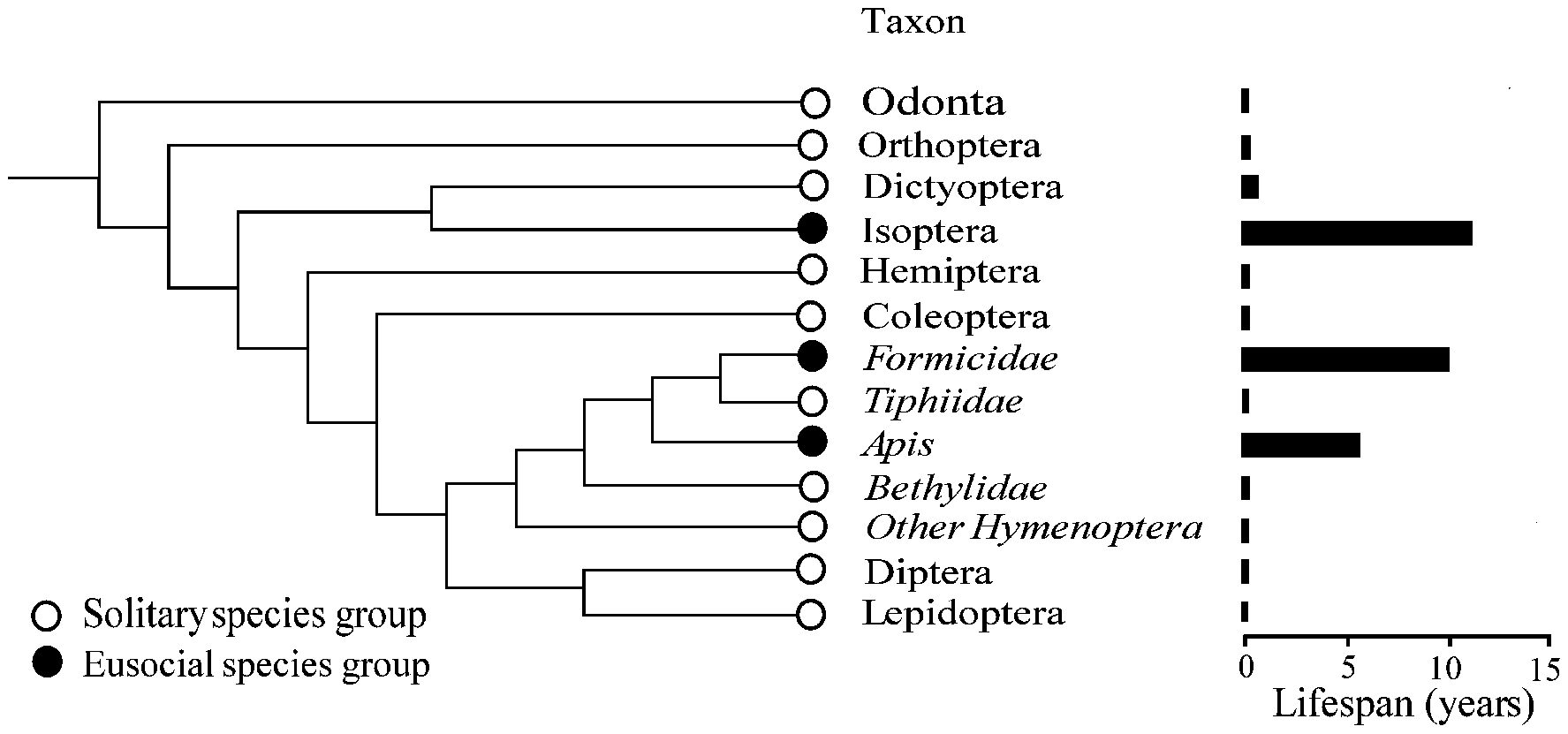 al. 2006, WOLSCHIN & AMDAM 2007a, b). Although con-firming fundamental differences between the subcastes in many basic pathways, these studies demonstrate the clas-sic limitation with proteomic and genomic aging studies. The typical experiment is to compare a long-lived and short-lived organism and the result is that the same basic meta-bolic pathways are usually affected. These include the in-sulin, TOR, and stress pathways. These are the three main biochemical processes that appear in many studies on aging
(BROUGHTON & PARTRIDGE 2009, GREWAL 2009, PAPA-CONSTANTINOU 2009, STANFEL & al. 2009), but also so
Fig. 1: The relative reproductive lifespan of various in-
fundamental to regulating metabolism that any major change
sect taxa showing the effect of eusociality. Details on the
in the organism's metabolism will likely perturb them. The
order Hymenoptera are given for several families because
important result is that the subcastes are different at a
eusociality evolved independently in the genus Apis and
basic metabolic level. This is a key lesson for anyone doing
in Formicidae (ants). Reprinted by permission from Mac-
an "omics" experiment on aging. Thus far the results have
millan Publishers Ltd: Nature (KELLER & GENOUD 1997),
been too general to be interpreted by themselves and have
proven most valuable as preliminary work for future re-
ductionistic studies (WHITFIELD & al. 2006, GRAFF & al.
istry of the size / lifespan trade-off in such eusocial sys-
2007). Most genomic studies require confirmatory work be-
tems given that the ant with the most genomic resources
fore meaningful results about specific genes and pathways
available (Solenopsis invicta) also has an extreme size poly-
morphism among its sterile workers (WANG & al. 2007,
One particularly relevant result for biomedicine is the
observed restoration of an age related decline in the im-mune system (AMDAM & al. 2004, 2005). It is well estab-
Social environment can change intrinsic
al. 2006, WOLSCHIN & AMDAM 2007a, b). Although con-firming fundamental differences between the subcastes in many basic pathways, these studies demonstrate the clas-sic limitation with proteomic and genomic aging studies. The typical experiment is to compare a long-lived and short-lived organism and the result is that the same basic meta-bolic pathways are usually affected. These include the in-sulin, TOR, and stress pathways. These are the three main biochemical processes that appear in many studies on aging
(BROUGHTON & PARTRIDGE 2009, GREWAL 2009, PAPA-CONSTANTINOU 2009, STANFEL & al. 2009), but also so
Fig. 1: The relative reproductive lifespan of various in-
fundamental to regulating metabolism that any major change
sect taxa showing the effect of eusociality. Details on the
in the organism's metabolism will likely perturb them. The
order Hymenoptera are given for several families because
important result is that the subcastes are different at a
eusociality evolved independently in the genus Apis and
basic metabolic level. This is a key lesson for anyone doing
in Formicidae (ants). Reprinted by permission from Mac-
an "omics" experiment on aging. Thus far the results have
millan Publishers Ltd: Nature (KELLER & GENOUD 1997),
been too general to be interpreted by themselves and have
proven most valuable as preliminary work for future re-
ductionistic studies (WHITFIELD & al. 2006, GRAFF & al.
istry of the size / lifespan trade-off in such eusocial sys-
2007). Most genomic studies require confirmatory work be-
tems given that the ant with the most genomic resources
fore meaningful results about specific genes and pathways
available (Solenopsis invicta) also has an extreme size poly-
morphism among its sterile workers (WANG & al. 2007,
One particularly relevant result for biomedicine is the
observed restoration of an age related decline in the im-mune system (AMDAM & al. 2004, 2005). It is well estab-
Social environment can change intrinsic