Practical medical trials have confirmed the maximum effect of Levitra, whith a positive effect being observed in 99 of 100 patients priligy uk Our internet store sells medicines produced in Indian pharmaceutical factories under licensed agreement.
Untitled
Acta Biochimica et Biophysica Sinica Advance Access published February 16, 2011
Acta Biochim Biophys Sin (2011): 1 – 10 | ª The Author 2011. Published by ABBS Editorial Office in association with Oxford University Press on behalf of the
Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences. DOI: 10.1093/abbs/gmr007.
PGC-1 coactivators in the control of energy metabolism
1Jiangsu Key Laboratory for Molecular and Medical Biotechnology, Nanjing Normal University, Nanjing 210046, China2Life Sciences Institute, University of Michigan, Ann Arbor, MI 48109, USA*Correspondence address. Tel: þ86-25-85891870 (C.L.)/þ1-734-615-3512 (J.L.); Fax: þ86-25-85891870(C.L.)/þ1-734-615-0495 (J.L.);E-mail: [email protected] (C.L.)/[email protected] (J.L.)
Chronic disruption of energy balance, where energy
in the year 2000 will carry a significant lifetime risk of
intake exceeds expenditure, is a major risk factor for the
developing type 2 diabetes, and therefore become prone to
development of metabolic syndrome. The latter is charac-
premature cardiovascular disease, blindness, kidney failure,
terized by a constellation of symptoms including obesity,
and amputations. A cardinal feature of metabolic syndrome
dyslipidemia, insulin resistance, hypertension, and non-
is severe obesity, which arises from chronic imbalance
alcoholic fatty liver disease. Altered expression of genes
between energy intake and energy consumption. As such,
involved in glucose and lipid metabolism as well as mito-
restoration of the energy balance is a major strategy for the
chondrial oxidative phosphorylation has been implicated
therapy of metabolic disease including obesity, diabetes,
in the pathogenesis of these disorders. The peroxisome
hypertension, atherosclerosis, and fatty liver diseases.
proliferator-activated receptor g coactivator-1 (PGC-1)
Peroxisome proliferator-activated receptor (PPAR) g
family of transcriptional coactivators is emerging as a
coactivator-1 (PGC-1) family members are multifunctional
hub linking nutritional and hormonal signals and energy
transcriptional coregulators that act as ‘molecular switches’
metabolism. PGC-1a and PGC-1b are highly responsive
in many metabolic pathways. PGC-1a and PGC-1b have
to environmental cues and coordinate metabolic gene pro-
been shown to regulate adaptive thermogenesis, mitochon-
grams through interaction with transcription factors and
drial biogenesis, glucose/fatty-acid metabolism, peripheral
chromatin-remodeling proteins. PGC-1a has been impli-
circadian clock, fiber-type switching in skeletal muscle,
cated in the pathogenic conditions including obesity,
and heart development. Their versatile actions are achieved
type 2 diabetes, neurodegeneration, and cardiomyopathy,
by interacting with different transcription factors in a
whereas PGC-1b plays an important role in plasma
tissue-specific manner. The potent effects of PGC-1 coacti-
lipoprotein homeostasis and serves as a hepatic target for
vators in coordinating various metabolic processes under-
niacin, a potent hypotriglyceridemic drug. Here, we
score their significant role in the control of energy
review recent advances in the identification of physiologi-
metabolism as well as their potential as targets for pharma-
cal and pathophysiological contexts involving PGC-1
coactivators, and also discuss their implications for thera-peutic development.
The PGC-1a gene is located on chromosome 5 in mice(chromosome 4 in humans) and encodes a protein contain-
ing 797 (mouse) or 798 (human) amino acids ]. Structural and functional studies have indicated thatPGC-1a has a strong transcriptional activation domain at
the N terminus, which interacts with several histone acetyl-transferase (HAT) complexes including 3’-5’-cyclic adeno-
The prevalence of contemporary life style, characterized by
sine monophosphate (cAMP) response element-binding
increased consumption of high-fat, high-fructose food and
protein (CREB)-binding protein, p300, and steroid receptor
reduced physical activity, has driven a dramatic increase in
coactivator-1 ]. These proteins acetylate histones and
the incidence of metabolic syndrome. It has been projected
remodel chromatin structure into a state that is permissive
that, by 2025, one in every three American children born
for transcriptional activation. Adjacent to the N-terminal
PGC-1 coactivators in the control of energy metabolism
domain is a regulatory region that roughly spans 200
energy demand, including the BAT, heart, skeletal muscle,
amino acids. Toward the C terminus, PGC-1a recruits the
kidney, and brain – In fact, when ectopically
thyroid receptor-associated protein/vitamin D receptor-
expressed in fat or muscle cells, PGC-1a strongly stimu-
interacting protein/mediator complex that facilitates direct
lates the program of nuclear and mitochondrial-encoded
interaction with the transcription initiation machinery [].
mitochondrial genes as well as organelle biogenesis [].
This region also interacts with the switch/sucrose non-
The stimulatory effects of PGC-1a on mitochondrial genes
fermentable (SWI/SNF) chromatin-remodeling complex
are achieved through its coactivation of nuclear respiratory
through its interaction with BAF60a The Ser/Arg-rich
factors 1 and 2 (NRF1 and NRF2, respectively) and the
domain and an RNA-binding domain toward C terminus
estrogen-related receptor a (ERRa) [The induc-
have been demonstrated to couple pre-mRNA splicing with
tion of NRF1 and NRF2 subsequently leads to the
transcription ]. As such, PGC-1a serves as a platform for
increased expression of mitochondrial transcription factor
the recruitment and assembly of various chromatin-
A (mtTFA) [as well as other mitochondrial subunits of
remodeling and histone-modifying enzymes to alter local
the electron transport chain complex such as b-adenosine-
chromatin state. Importantly, the PGC-1a transcriptional
triphosphate (ATP) synthase, cytochrome c, and cyto-
activator complex is also able to displace repressor pro-
chrome oxidase IV []. mtTFA translocates to mito-
teins, such as histone deacetylase and small heterodimer
chondrial matrix, where it stimulates mitochondrial DNA
partner, on its target promoters, providing an alternative
replication and mitochondrial gene expression
mechanism for gene activation [PGC-1a and PGC-1b
As mentioned above, a critical aspect of PGC-1a is
share extensive domain similarity and several clusters of
that it is highly versatile and has the ability to increase
conserved amino acids, such as the LXXLL motif that
the transcriptional activity of many nuclear receptor
interacts with nuclear receptors and host cell factor 1 inter-
families, including members of the estrogen, PPAR, reti-
noid X, mineralocorticoid, glucocorticoid (GR), liver X
(PGC-1-related coactivator), also contains the activation
(LXR), pregnane X, the constitutive androstane, vitamin
domain and RNA-binding domain, but overall has more
limited homology to PGC-1a and PGC-1b []. The PGC-1
PGC-1a can also bind to unliganded nuclear receptors,
family members are conserved in higher vertebrates,
as in the case of the orphan hepatic nuclear factor
including mammals, birds, and fish. Interestingly, a PGC-1
(HNF) 4a, farnesoid X receptor (FXR), and ERRa,
family homologue named Spargel was recently identified
suggesting that their conformations are conductive to
in Drosophila that could regulate mitochondrial activity
ligand-independent mechanisms of gene regulation [].
PGC-1a transcriptional partners are not limited to the
Both PGC-1a and PGC-1b robustly regulate mitochon-
nuclear receptor superfamily; however, this coactivator
drial oxidative metabolism (Fig. ). PGC-1a was initially
also associates with a diverse array of other transcription
identified as a PPARg-interacting protein from the brown
factors, including forkhead/winged helix protein family
adipose tissue (BAT) that could regulate adaptive thermo-
member FOXO1, as well as a number of zinc-finger pro-
genesis in response to cold []. Subsequent studies
revealed that the core function of PGC-1a was to stimulate
screen ]. The docking interface for these interacting
proteins appears to distribute throughout the length of
PGC-1a is abundantly expressed in tissues with high
PGC-1a. In addition, PGC-1a has three functional
Figure 1 The working model of PGC-1 coactivators
PGC-1a and PGC-1b regulate diverse metabolic programs through coactivating selective
transcriptional factors (TF) associated with regulatory elements of target genes. PGC-1 recruits HAT, SWI/SNF chromatin-remodeling, Sirt1 deacetylase,and mediator complexes to modulate the epigenetic status of chromatin.
PGC-1 coactivators in the control of energy metabolism
LXXLL motifs that are responsible for docking nuclear
PGC-1a function by repressing its transcription [].
Impaired PGC-1a expression and mitochondrial function
proteins enables PGC-1a to regulate various metabolic
contributes to neurodegeneration in susceptible neurons
processes in a tissue-specific manner.
In addition, PGC-1a plays an important role in theregulation of genes responsible for the detoxification of
Tissue-specific metabolic actions of PGC-1
reactive oxygen species (ROS), including copper/zinc
superoxide dismutase (SOD1), manganese SOD (SOD2),and glutathione peroxidase 1 ]. In this case, PGC-1a
The following section reviews PGC-1 functions in oxi-
protects dopaminergic neurons from degeneration caused
dative tissues including the brain, heart, brown fat, skeletal
by oxidative stress. Taken together, the finding that
muscle, liver, and pancreatic islets, based on gain and
PGC-1a expression is impaired in the striatum of HD
loss-of-function analysis both in cultured cells and in vivo.
patients raises the possibility that molecules activating
A summary of tissue-specific PGC-1 functions and the
PGC-1a may be therapeutically useful.
phenotype of PGC-1 transgenic mouse models are includedin and respectively.
Heart is an organ with an extremely high and dynamic
demand for ATP. Much of this supply comes from
fatty-acid b-oxidation, though glucose also serves as fuel
several brain areas, predominantly in the striatum, and
source. Several studies have demonstrated that PGC-1a is a
exhibit behavioral abnormalities including marked hyper-
crucial regulator of oxidative metabolism in the heart.
activity and frequent limb clasping Recent studies
PGC-1a mRNA levels are strongly induced in the neonatal
heart, along with the activation of mitochondrial biogenesis
support a crucial role of this factor in neuronal function
and the metabolic switch from glycolysis to oxidative
and energy balance. Similar brain lesions are observed
phosphorylation in cardiac muscle ]. Overexpression of
when PGC-1a is selectively ablated in CaMKIIa-positive
PGC-1a both in vitro and in vivo powerfully induces mito-
neurons, providing direct evidence for its action in
chondrial gene expression and biogenesis ]. PGC-1a
neurons. Of note, striatal degeneration with hyperactivity
expression is reduced in several animal models of cardiac
is reminiscent of Huntington’s disease (HD) in humans,
dysfunction, which is typically accompanied by a meta-
potentially implicating PGC-1a in the selective vulner-
bolic switch from fat oxidation to glycolysis []. PGC-1a
ability of striatal neurons in HD. To date, the specific
null mice exhibit significantly lower cardiac reserve in
role of PGC-1a in linking mitochondrial dysfunction to
HD pathogenesis has been explored. The mutant hun-
Moreover, PGC-1a null mice develop early symptoms of
tingtin protein accumulated in HD brain interferes with
heart failure, such as activation of the fetal program of
Table 1 Tissue-specific functions of PGC-1a and PGC-1b
Maintenance of mitochondrial function ROS detoxification
Mitochondrial oxidative metabolism [], fatty-acid b-oxidation
Mitochondrial OXPHOS mediating the effects of
Mitochondrial biogenesis and fat oxidation [Adaptive
Brown adipocyte differentiation []; Adaptive
Slow-switch muscle fiber [], mitochondrial biogenesis
Hepatic fasting response [], homocysteine metabolism [],
integration of circadian clock and metabolism []
Suppression of GSIS and membrane depolarization []
PGC-1 coactivators in the control of energy metabolism
Table 2 Phenotypes of PGC-1 knockout and transgenic mouse models
Hyperactive, cold-sensitive, resistant to diet-induced obesity, lesions in the
Reduced muscle performance and exercise capacity, impaired adaptive
Impaired glucose tolerance, normal peripheral insulin sensitivity
Switch of type II muscle fiber to type IIa and I muscle fibers, resistance to
Loss of sarcomeric structure, dilated cardiomyopathy
Increased mitochondrial biogenesis, derangements of mitochondrial ultrastructure,
Impaired mitochondrial function, reduced body weight and fat mass, increased
thermogenesis, blunted chronotropic response to dobutamine in the heart, hepatic
steatosis, reduced lipoprotein-associated triglyceride and cholesterol content
Decreased activity during the dark cycle, abnormal hypothermia and morbidity,
hepatic steatosis, increased serum triglyceride and cholesterol
Mitochondrial dysfunction in liver and skeletal muscle, hepatic steatosis, hepatic
Increased fatty-acid oxidation, hyperphagia, reduced body weight and adipose
tissue, increased exercise capacity, increased IIX fiber content
Neonatal lethality, bradycardia, intermittent heart block, reduced cardiac output,
reduced growth, a late fetal arrest in mitochondrial biogenesis
cardiac gene expression and a significant increase in circu-
lating levels of atrial natriuretic peptide, a hallmark of
In rodents, BAT is the major organ responsible for adaptive
cardiac dysfunction These mice also exhibit lower
thermogenesis during cold exposure. In contrast to white
treadmill-running capacity and diminished cardiac function
adipose tissue, whose primary physiological function is
after exercise. However, it should be noted that superphy-
energy storage, the main function of BAT is energy dissi-
siological expression of PGC-1a in the heart leads to
pation, largely in the form of heat. The expression of
robust mitochondrial proliferation and myofibrillar displa-
PGC-1a is strongly induced in BAT by cold temperature;
cement, and dilated cardiomyopathy ensues [As such,
PGC-1a is downstream of the b-adrenergic receptor/cAMP
therapeutic regulation of PGC-1a in heart failure should
pathway and sympathetic nervous system activity ].
aim at restoring PGC-1a function in cardiac muscle within
In this case, PGC-1a turns on several key components
a therapeutically beneficial window.
involved in the adaptive thermogenic program, including
PGC-1b is also abundantly expressed in the heart [].
the stimulation of fuel uptake, mitochondrial fatty-acid
Heart function in PGC-1b-deficient mice is largely unaf-
b-oxidation, and stimulation of uncoupling protein 1
fected under normal conditions []. However, PGC-1b
(UCP1) expression ]. PGC-1a interacts with other
ablation reduces mitochondrial content in cardiac muscle
nuclear hormone receptors such as PPARa, retinoic acid
and blunts the effect of adrenergic stimulation on heart rate
receptor, and thyroid receptor to enhance UCP1 expression.
Remarkably, mice with combined deficiency of
PGC-1a and PGC-1b (PGC-1ab2/2) die shortly after birth
generating heat and uncouples oxidative phosphorylation
with small hearts, bradycardia, intermittent heart block, and
from ATP production. PGC-1a-deficient mice are unable
a markedly reduced cardiac output Cardiac-specific
to defend against cold stress due to thermogenic defects
ablation of PGC-1b on a PGC-1a-deficient background
results in cardiac defects including reduced growth, a late
PGC-1b mRNA is induced during white and brown
fetal arrest in mitochondrial biogenesis, and persistence of a
adipocyte differentiation ]. Interestingly, while the
fetal pattern of gene expression These observations
expression of PGC-1b is not cold inducible, its deficiency
suggest that PGC-1a and PGC-1b collectively are required
also impairs adaptive thermogenesis [], suggesting that
for the postnatal metabolic and functional maturation of the
these two coactivators play non-redundant function in fuel
PGC-1 coactivators in the control of energy metabolism
RNAi-mediated liver-specific PGC-1a knockdown mice
PGC-1a is abundantly expressed in skeletal muscle, par-
display the impairment of gluconeogenic gene expression
ticularly slow-twitch myofibers, and is rapidly inducible by
and hepatic glucose production []. These mice
exercise training in rodents and humans – It is
develop hypoglycemia and hepatic steatosis upon fasting
clear that calcium signaling pathways play important roles
In addition, PGC-1a regulates the genes encoding
in the induction of PGC-1a through calcineurin and
homocysteine synthesis enzymes in the liver and modulates
calcium-dependent protein kinases and the subsequent acti-
plasma homocysteine levels ]. Forced expression of
vation of CREB and myocyte-enhancing factor 2 ].
PGC-1a in vivo leads to elevated plasma homocysteine
In addition, p38 mitogen-activated protein kinase ( p38
MAPK) and AMP-dependent kinase (AMPK) are also
In mammals, circadian clock regulates major aspects of
required for exercise-induced PGC-1a expression ].
energy metabolism, including glucose and lipid homeosta-
Interestingly, muscle-specific overexpression muscle cre-
sis and mitochondrial respiration. Our recent work revealed
atine kinase (MCK) of PGC-1a in mice turns white, glyco-
that PGC-1a is a key component of the circadian oscillator
lytic skeletal muscles (fast-twitch muscle fibers) into red,
that integrates the peripheral clock and energy metabolism
PGC-1a stimulates the expression of Bmal1, a core
clock gene, in hepatocytes and muscle cells through coacti-
vation of the ROR family of orphan nuclear receptors.
slow-twitch myofibers ]. In addition to the regulation of
Mice lacking PGC-1a have abnormal diurnal rhythms of
mitochondrial function, PGC-1a increases mRNA content
activity, body temperature, and metabolic rate. As PGC-1a
of enzymes involved in fat metabolism such as fatty-acid
expression is regulated by nutritional and hormonal cues, it
translocase/CD36, carnitine palmitoyltransferase I, and
is likely that it links these signals to the clockwork and
medium-chain acyl-coenzyme A dehydrogenase (MCAD)
synchronizes tissue metabolism with circadian pacemaker.
in skeletal muscle [Consistent with the molecular
PGC-1b expression is increased in response to dietary
changes, PGC-1a transgenic muscle has increased fatigue
intake of fats and leads hyperlipidemia through activating
resistance following electrical stimulation [In contrast,
both whole-body and muscle-specific PGC-1a knock out
(VLDL) secretion []. Several factors are involved in med-
(KO) mice show reduced mRNA and/or protein content of
iating the effects of PGC-1b on plasma triglyceride metab-
mitochondrial respiratory chain proteins and ATP synthase.
olism, including sterol response element-binding protein
They are exercise intolerant and their skeletal muscles are
(SREBP), LXR, and Foxa2 Recent studies demon-
strated that PGC-1b and its target gene apolipoprotein C3
In primary cultures of rat muscle cells, PGC-1b
(ApoC3) are downstream of nicotinic acid, a widely used
increases the expression of glucose transporter 4, myosin
hypotriglyceridemic drug []. Both acute injection and
heavy chain Ib, and other slow-twitch muscle markers
chronic feeding of mice with nicotinic acid suppress
While PGC-1b also stimulates mitochondrial biogen-
PGC-1b and ApoC3 expression in the liver [These
esis in skeletal muscle, it appears to drive a program of
studies illustrated a new role for PGC-1b in modulating
gene expression that is reminiscent of type IIx fibers [].
lipoprotein catabolism and the relevance of this pathway in
In addition, the expression of PGC-1b, but not PGC-1a, is
therapeutic action of nicotinic acid. Remarkably, systemic
decreased along with reduced ERRa activity and MCAD
delivery of antisense oligonucleotide targeting PGC-1b
expression in skeletal muscle of senescence-accelerated
improved systemic metabolic homeostasis in the model of
fructose-induced insulin resistance ].
Hepatic PGC-1a expression reaches its peak during early
b-Cell dysfunction is cardinal for the development of type
postnatal period []. In adults, starvation induces PGC-1a
2 diabetes. PGC-1a is expressed in pancreatic b-cells and
expression in the liver through glucagon and GR signaling
is elevated in animal models of type 2 diabetes [].
PGC-1a orchestrates a complex program of metabolic
Ectopic expression of PGC-1a impairs both early and
changes that occur during the transition of a fed to a fasted
delayed glucose-stimulated insulin secretion (GSIS) and
liver, including gluconeogenesis, fatty-acid b-oxidation,
suppresses membrane depolarization without affecting
ketogenesis, heme biosynthesis, and bile-acid homeostasis.
baseline insulin secretion. Altered PGC-1a expression is
These effects of PGC-1a on fasting adaption are achieved
accompanied by increased glucose-6-phosphatase and
by coactivating key hepatic transcription factors, such as
reduced glucokinase gene expression. Furthermore, UCP2
HNF4a, PPARa, GR, FOXO1, FXR, and LXR []. In
may be another effector downstream of PGC-1; UCP2
accordance with these observations, PGC-1a KO mice and
PGC-1 coactivators in the control of energy metabolism
production, and negatively regulates GSIS Although
the mechanism through which PGC-1a is induced in dia-betic animal models is not understood, fatty acids and
As PGC-1a regulates multiple aspects of energy metab-
incomplete inactivation of FOXO1 may contribute to
olism, it is not surprising that PGC-1a has been found to
this process. In contrast to animal data, studies in human
be dysregulated in several pathological conditions. The
type 2 diabetic islets showed that PGC-1a mRNA
expression of PGC-1a and its target genes involved in
expression is markedly reduced and correlated with the
mitochondrial oxidative phosphorylation (OXPHOS) is sig-
reduction in insulin secretion in those islets ]. DNA
nificantly decreased in the skeletal muscle of patients with
methylation of the PGC-1a gene promoter is increased in
type 2 diabetes ]. Similar reduction of PGC-1a
human diabetic islets. Therefore, the exact function of
expression was also observed in the adipose tissue of
PGC-1a in pancreatic islets needs further study.
insulin-resistant and morbidly obese individuals [].
The function of PGC-1b in islets is less studied. A
Interestingly, thiozolidinedione, an important class of anti-
recent study indicated that PGC-1b, in contrast to PGC-1a,
diabetic drugs, can enhance the expression of PGC-1a and
directly binds to and acts as a coactivator of SREBPs and
mitochondrial biogenesis in white adipose tissue [].
Foxa2 involved in pancreas development and function
While these observations support a potentially beneficial
The authors also showed that PGC-1b suppresses
role of PGC-1a in insulin resistance and type 2 diabetes,
GSIS via upregulation of UCP2 and granuphilin gene
several studies suggested distinct actions of PGC-1a in
other tissues. For example, PGC-1a expression is elevatedin the liver of both type 1 and type 2 diabetic mousemodels ]. Furthermore, PGC-1a has been shown to
stimulate hepatic glucose production and suppress b-cell
Post-translational modifications of PGC-1
energy metabolism and insulin release in mice [].
Paradoxically, transgenic expression of PGC-1a in skeletalmuscle leads to robust mitochondrial biogenesis but also
PGC-1a undergoes extensive post-translational modifi-
causes insulin resistance, likely the result of imbalance of
cations, including acetylation, phosphorylation, methyl-
lipid uptake and oxidation [In addition, a common
ation, and SUMOylation, in response to nutritional and
polymorphism of the PGC-1a gene (Gly482Ser), which
hormonal signals. These modifications allow fine-tuning of
apparently reduces PGC-1a activity, has been linked to
PGC-1a activities in a context-dependent manner. The
acetyl transferase general control of amino-acid synthesis
In the cardiovascular system, PGC-1a expression is also
5 acetylates PGC-1a at several lysine residues, alters its
decreased in hypertrophic heart ]. PGC-1a null mice
localization within the nucleus, and inhibits its transcrip-
display accelerated cardiac dysfunction and clinical signs
tional activity On the contrary, deacetylation of
of heart failure In contrast, PPARa ligand-dependent
PGC-1a through sirtuin 1 (Sirt1) increases PGC-1a activity
transcriptional activity and coactivation by PGC-1a are
on gluconeogenic gene transcription in the liver [].
enhanced in the heart by stress including ischemia and
PGC-1a is phosphorylated by both p38 MAPK and AMPK
hypoxia ]. In peripheral vessel tissues, downregulation
in skeletal muscle [], leading to a more stable and
of PGC-1a expression was observed in vascular smooth
active protein. In contrast, phosphorylation of PGC-1a by
muscle cells (VSMCs) treated by oleic acid and high
Akt/protein kinase B downstream of the insulin signaling
glucose []. Restoration of PGC-1a has beneficial effects
cascade in the liver decreased its stability and transcriptional
on VSMCs and endothelial cells ]. In this context,
activity []. In addition, PGC-1a also undergoes methyl-
PGC-1a appears to play an important role in ROS metab-
ation at several arginine residues in the C-terminal region
olism and defense against oxidative stress. These obser-
by protein arginine methyltransferase 1 ]. Finally,
vations indicate that PGC-1a is an important factor in the
PGC-1a can undergo SUMOylation in conserved lysine
regulation of cardiovascular function.
residue 183 and its transcriptional activity is attenuated
Abnormalities in mitochondrial function are associated
Interestingly, experiments using C2C12 cells have indi-
with neurodegenerative disorders including Parkinson’s
disease, Alzheimer’s disease, and HD. Levels of PGC-1a
PGC-1a for deacetylation by Sirt1 [], suggesting that
are reduced in the brain of HD patients due to repression
different modifications of PGC-1a likely communicate with
of PGC-1a gene expression by mutant huntingtin, leading
each other to coordinately regulate its activity. PGC-1b is
to mitochondrial defects and increased oxidative stress
also acetylated at multiple sites however, the biologi-
Expression of PGC-1a partially reverses the toxic
cal significance of these events is less well defined.
effects and provides neuroprotection in the HD mutant
PGC-1 coactivators in the control of energy metabolism
mouse ]. In the peripheral nervous system, PGC-1a has
clinical use. For example, coactivators have kinetic benefits
been shown to regulate gene expression at the neuromuscu-
in controlling biological programs in that they coordinate
lar junction and influences expression of acetylcholine
different steps in biological programs through the inte-
receptors in muscle fibers In addition, elevated
gration of the activity of various transcription factors. As
PGC-1a levels protect neural cells in culture from cell
such, significant biological effects can be achieved by
death caused by oxidative-stressor through its induction of
quantitatively modulating coactivator function. The diver-
sity of post-translational modifications of PGC-1 poten-
Energy metabolism in cancer cells differs fundamentally
tially allows targeting specific protein – protein interaction
from that in its normal counterparts. In general, cancer
interface. As discussed above, tissue-specific modulation of
cells have high glycolytic activity and prefer glucose as a
PGC-1 function is essential for metabolic modulation
fuel source, a phenomenon known as the Walburg effect.
without causing deleterious side effects.
The switch from OXPHOS to glycolysis occurs even in thepresence of sufficient oxygen. This aerobic glycolysis has
been postulated to enhance cancer cell proliferationand survival. Interestingly, reduced expression of PGC-1a
Owing to space limitations, we apologize to those whose
has been observed in human breast cancer colon
publications related to the discussed issues could not
cancer [], liver cancer and ovarian cancer [].
Adenoviral-mediated overexpression of PGC-1a inducesE-cadherin expression while decreasing motility of humanhepatoma HepG2 cells Such manipulation also causes
cell apoptosis in human ovarian cancer cells through aPPARg-dependent pathway These findings suggest
This work was supported by the grants from the National
that PGC-1a is a potentially important regulator of cancer
Natural Science Foundation of China (30870928), the
cell metabolism and contributes to altered metabolic
Research Fund for the Doctoral Program of Higher
function in cancer cells. A causal relationship between
Education of China (20103207110007), the Fok Ying
PGC-1a and cancer development, however, remains to be
Tong Education Foundation (121022), the Major Program
(09KJA180004) (to C.L.), and the NIH grant (DK077086)
The PGC-1 family of transcriptional coactivators has
emerged as a regulatory hub within the transcriptional net-works that maintain metabolic homeostasis. The dynamic
1 Puigserver P, Wu Z, Park CW, Graves R, Wright M and Spiegelman BM.
regulation of PGC-1 expression and/or post-translational
A cold-inducible coactivator of nuclear receptors linked to adaptive ther-
modification in response to nutritional and hormonal
mogenesis. Cell 1998, 92: 829 – 839.
signals provides a highly versatile regulatory platform for
2 Puigserver P, Adelmant G, Wu Z, Fan M, Xu J, O’Malley B and
metabolic control. A major challenge is to map out tissue-
Spiegelman BM. Activation of PPARg coactivator-1 through transcription
specific activities of PGC-1a and PGC-1b as well as the
factor docking. Science 1999, 286: 1368 – 1371.
transcriptional partners that mediate PGC-1 actions. Of
3 Wallberg AE, Yamamura S, Malik S, Spiegelman BM and Roeder RG.
Coordination of p300-mediated chromatin remodeling and TRAP/mediator
note, recent genome-wide coactivation analyses provide a
function through coactivator PGC-1a. Mol Cell 2003, 12: 1137 – 1149.
global view of potential PGC-1a interacting proteins in
4 Li S, Liu C, Li N, Hao T, Han T, Hill DE and Vidal M, et al.
humans. While the biological function of nuclear receptor
Genome-wide coactivation analysis of PGC-1a identifies BAF60a as a
targets of PGC-1 is better understood, very little is known
regulator of hepatic lipid metabolism. Cell Metab 2008, 8:105 – 117.
about a large number of zinc-finger proteins that associate
5 Monsalve M, Wu Z, Adelmant G, Puigserver P, Fan M and Spiegelman
BM. Direct coupling of transcription and mRNA processing through the
with PGC-1a. The identification of PGC-1a splicing iso-
thermogenic coactivator PGC-1. Mol Cell 2000, 6: 307 – 316.
forms also adds additional complexity Dysregulation
6 Borgius LJ, Steffensen KR, Gustafsson JA and Treuter E. Glucocorticoid
of PGC-1 expression has been observed in a wide variety
signaling is perturbed by the atypical orphan receptor and core pressor
of pathological conditions. As such, proper modulation of
SHP. J Biol Chem 2002, 277: 49761 – 49766.
PGC-1a expression/activity has great potential in the
7 Lin J, Tarr PT, Yang R, Rhee J, Puigserver P, Newgard CB and
prevention and treatment of diseases associated with
Spiegelman BM. PGC-1b in the regulation of hepatic glucose and energy
metabolism. J Biol Chem 2003, 278: 30843 – 30848.
impaired mitochondrial function and oxidative metabolism.
8 Andersson U and Scarpulla RC. Pgc-1-related coactivator, a novel,
Although targeting a coactivator could be challenging,
serum-inducible coactivator of nuclear respiratory factor 1-dependent tran-
certain features of coactivator action can be exploited for
scription in mammalian cells. Mol Cell Biol 2001, 21: 3738 – 3749.
PGC-1 coactivators in the control of energy metabolism
9 Tiefenbo¨ck SK, Baltzer C, Egli NA and Frei C. The Drosophila PGC-1
27 Uldry M, Yang W, St-Pierre J, Lin J, Seale P and Spiegelman BM.
homologue Spargel coordinates mitochondrial activity to insulin signaling.
Complementary action of the PGC-1 coactivators in mitochondrial biogen-
esis and brown fat differentiation. Cell Metab 2006, 3: 333 – 341.
10 Wu Z, Puigserver P, Andersson U, Zhang C, Adelmant G, Mootha V and
28 Lelliott CJ, Medina-Gomez G, Petrovic N, Kis A, Feldmann HM, Bjursell
Troy A, et al. Mechanisms controlling mitochondrial biogenesis and res-
M and Parker N, et al. Ablation of PGC-1b results in defective mitochon-
piration through the thermogenic coactivator PGC-1. Cell 1999, 98:
drial activity, thermogenesis, hepatic function, and cardiac performance.
11 Esterbauer H, Oberkofler H, Krempler F and Patsch W. Human peroxi-
29 Lin J, Wu H, Tarr PT, Zhang CY, Wu Z, Boss O and Michael LF, et al.
some proliferator activated receptor g coactivator 1 (PPARGC1) gene:
Transcriptional coactivator PGC-1a drives the formation of slow-twitch
cDNA sequence, genomic organization, chromosomal localization, and
muscle fibres. Nature 2002, 418: 797 – 801.
tissue expression. Genomics 1999, 62: 98 – 102.
30 Miura S, Tomitsuka E, Kamei Y, Yamazaki T, Kai Y, Tamura M and Kita
12 Knutti D, Kaul A and Kralli A. A tissue-specific coactivator of steroid
K, et al. Overexpression of PGC-1a leads to muscle atrophy with
receptors, identified in a functional genetic screen. Mol Cell Biol 2000,
depletion of ATP. Am J Pathol 2006, 169: 1129 – 1139.
31 Arany Z, Lebrasseur N, Morris C, Smith E, Yang W, Ma Y and Chin
13 Mootha VK, Handschin C, Arlow D, Xie X, St Pierre J, Sihag S and
S, et al. The transcriptional coactivator PGC-1b drives the formation of
Yang W, et al. Erra and Gabpa/b specify PGC-1a-dependent oxidative
oxidative type IIX fibers in skeletal muscle. Cell Metab 2007, 5:
phosphorylation gene expression that is altered in diabetic muscle. Proc
Natl Acad Sci USA 2004, 101: 6570 – 6575.
32 Yoon JC, Puigserver P, Chen G, Donovan J, Wu Z, Rhee J and Adelmant
14 Schreiber SN, Emter R, Hock MB, Knutti D, Cardenas J, Podvinec M and
G, et al. Control of hepatic gluconeogenesis through the transcriptional
Oakeley EJ, et al. The estrogen-related receptor a (ERRa) functions in
coactivator PGC-1. Nature 2001, 413: 131 – 138.
PPARg coactivator 1a (PGC-1a)-induced mitochondrial biogenesis. Proc
33 Li S, Arning E, Liu C, Vitvitsky V, Hernandez C, Banerjee R and
Natl Acad Sci USA 2004, 101: 6472 – 6477.
Bottiglieri T, et al. Regulation of homocysteine homeostasis through the
15 Scarpulla RC. Nuclear activators and coactivators in mammalian mito-
transcriptional coactivator PGC-1a. Am J Physiol Endocrinol Metab 2009,
chondrial biogenesis. Biochim Biophys Acta 2002, 1576: 1 – 14.
16 Scarpulla RC. Transcriptional activators and coactivators in the nuclear
34 Liu C, Li S, Liu T, Borjigin J and Lin JD. Transcriptional coactivator
control of mitochondrial function in mammalian cells. Gene 2002, 286:
PGC-1a integrates the mammalian clock and energy metabolism. Nature
17 Larsson NG, Wang J, Wilhelmsson H, Oldfors A, Rustin P, Lewandoski
35 Lin J, Yang R, Tarr PT, Wu PH, Handschin C, Li S and Yang W, et al.
M and Barsh GS, et al. Mitochondrial transcription factor A is necessary
Hyperlipidemic effects of dietary saturated fats mediated through PGC-1b
for mtDNA maintenance and embryogenesis in mice. Nat Genet 1998, 18:
coactivation of SREBP. Cell 2005, 120: 261 – 273.
36 Hernandez C, Molusky M, Li Y, Li S and Lin JD. Regulation of hepatic
18 Lin J, Handschin C and Spiegelman BM. Metabolic control through
ApoC3 expression by PGC-1b mediates hypolipidemic effect of nicotinic
the PGC-1 family of transcription coactivators. Cell Metab 2005, 1:
acid. Cell Metab 2010, 12: 411 – 419.
37 Yoon JC, Xu G, Deeney JT, Yang SN, Rhee J, Puigserver P and Levens
19 Liang H and Ward WF. PGC-1a: a key regulator of energy metabolism.
AR, et al. Suppression of b cell energy metabolism and insulin release by
Adv Physiol Educ 2006, 30: 145 – 151.
PGC-1a. Dev Cell 2003, 5: 73 – 83.
20 Soyal S, Krempler F, Oberkofler H and Patsch W. PGC-1a: a potent tran-
38 Oberkofler H, Hafner M, Felder T, Krempler F and Patsch W.
scriptional cofactor involved in the pathogenesis of type 2 diabetes.
Diabetologia 2006, 49: 1477 – 1488.
(PPAR) g coactivator-1b is involved in the regulation of glucose-stimulated
21 Weydt P, Pineda VV, Torrence AE, Libby RT, Satterfield TF, Lazarowski
insulin secretion in INS-1E cells. J Mol Med 2009, 87: 299 – 306.
ER and Gilbert ML, et al. Thermoregulatory and metabolic defects in
39 Lin J, Wu PH, Tarr PT, Lindenberg KS, St-Pierre J, Zhang CY and
Huntington’s disease transgenic mice implicate PGC-1a in Huntington’s
Mootha VK, et al. Defects in adaptive energy metabolism with
disease neurodegeneration. Cell Metab 2006, 4: 349 – 362.
CNS-linked hyperactivity in PGC-1a null mice. Cell 2004, 119: 121 – 135.
22 St-Pierre J, Drori S, Uldry M, Silvaggi JM, Rhee J, Ja¨ger S and
40 Leone TC, Lehman JJ, Finck BN, Schaeffer PJ, Wende AR, Boudina S
Handschin C, et al. Suppression of reactive oxygen species and neurode-
and Courtois M, et al. PGC-1a deficiency causes multisystem energy
generation by the PGC-1 transcriptional coactivators. Cell 2006, 127:
metabolic derangements: muscle dysfunction, abnormal weight control and
hepatic steatosis. PLoS Biol 2005, 3: e101.
23 Cowell RM, Blake KR and Russell JW. Localization of the transcriptional
41 Handschin C, Choi CS, Chin S, Kim S, Kawamori D, Kurpad AJ and
coactivator PGC-1a to GABAergic neurons during maturation of the rat
Neubauer N, et al. Abnormal glucose homeostasis in skeletal muscle-
brain. J Comp Neurol 2007, 502: 1 – 18.
specific PGC-1a knockout mice reveals skeletal muscle-pancreatic beta
24 Lehman JJ, Barger PM, Kovacs A, Saffitz JE, Medeiros DM and Kelly
cell crosstalk. J Clin Invest 2007, 117: 3463 – 3474.
42 Russell LK, Mansfield CM, Lehman JJ, Kovacs A, Courtois M, Saffitz JE
promotes cardiac mitochondrial biogenesis. J Clin Invest 2000, 106:
and Medeiros DM, et al. Cardiac-specific induction of the transcriptional
coactivator PGC-1a promotes mitochondrial biogenesis and reversible car-
25 Lai L, Leone TC, Zechner C, Schaeffer PJ, Kelly SM, Flanagan DP and
diomyopathy in a developmental stage-dependent manner. Circ Res 2004,
Medeiros DM, et al. Transcriptional coactivators PGC-1a and PGC-1b
control overlapping programs required for perinatal maturation of the heart.
43 Vianna CR, Huntgeburth M, Coppari R, Choi CS, Lin J, Krauss S and
Barbatelli G, et al. Hypomorphic mutation of PGC-1b causes mitochon-
26 Sonoda J, Mehl IR, Chong LW, Nofsinger RR and Evans RM. PGC-1b
drial dysfunction and liver insulin resistance. Cell Metab 2006, 4:
controls mitochondrial metabolism to modulate circadian activity, adaptive
thermogenesis, and hepatic steatosis. Proc Natl Acad Sci USA 2007, 104:
44 Cui L, Jeong H, Borovecki F, Parkhurst CN, Tanese N and Krainc D.
Transcriptional repression of PGC-1a by mutant huntingtin leads to
PGC-1 coactivators in the control of energy metabolism
mitochondrial dysfunction and neurodegeneration. Cell 2006, 127:
signaling to Pdx1 regulation of pancreatic b cell growth. J Clin Invest
45 Huss JM and Kelly DP. Mitochondrial energy metabolism in heart failure:
63 Ling C, Del Guerra S, Lupi R, Ro¨nn T, Granhall C, Luthman H and
a question of balance. J Clin Invest 2005, 115: 547 – 555.
Masiello P, et al. Epigenetic regulation of PPARGC1A in human type 2
46 Arany Z, He H, Lin J, Hoyer K, Handschin C, Toka O and Ahmad F,
diabetic islets and effect on insulin secretion. Diabetologia 2008, 51:
et al. Transcriptional coactivator PGC-1a controls the energy state and
contractile function of cardiac muscle. Cell Metab 2005, 1: 259 – 271.
64 Lerin C, Rodgers JT, Kalume DE, Kim SH, Pandey A and Puigserver P.
47 Lin J, Puigserver P, Donovan J, Tarr P and Spiegelman BM. Peroxisome
GCN5 acetyltransferase complex controls glucose metabolism through
proliferator-activated receptor gamma coactivator 1b (PGC-1b), a novel
transcriptional repression of PGC-1a. Cell Metab 2006, 3: 429 – 438.
PGC-1-related transcription coactivator associated with host cell factor. J
65 Rodgers JT, Lerin C, Haas W, Gygi SP, Spiegelman BM and Puigserver P.
Nutrient control of glucose homeostasis through a complex of PGC-1a
48 Baar K, Wende AR, Jones TE, Marison M, Nolte LA, Chen M and Kelly
and SIRT1. Nature 2005, 434: 113 – 118.
DP, et al. Adaptations of skeletal muscle to exercise: rapid increase in the
66 Puigserver P, Rhee J, Lin J, Wu Z, Yoon JC, Zhang CY and Krauss S,
transcriptional coactivator PGC-1. FASEB J 2002, 16: 1879 – 1886.
49 Goto M, Terada S, Kato M, Katoh M, Yokozeki T, Tabata I and
MAP kinase activation of PPARg coactivator-1. Mol Cell 2001, 8:
Shimokawa T. cDNA cloning and mRNA analysis of PGC-1 in epitro-
chlearis muscle in swimming-exercised rats. Biochem Biophys Res
67 Jager S, Handschin C, St-Pierre J and Spiegelman BM. AMP-activated
protein kinase (AMPK) action in skeletal muscle via direct phosphoryl-
50 Norrbom J, Sundberg CJ, Ameln H, Kraus WE, Jansson E and Gustafsson
ation of PGC-1a. Proc Natl Acad Sci USA 2007, 104: 12017 – 12022.
T. PGC-1a mRNA expression is influenced by metabolic perturbation in
68 Li X, Monks B, Ge Q and Birnbaum MJ. Akt/PKB regulates hepatic
exercising human skeletal muscle. J Appl Physiol 2004, 96: 189 – 194.
metabolism by directly inhibiting PGC-1a transcription coactivator. Nature
51 Akimoto T, Pohnert SC, Li P, Zhang M, Gumbs C, Rosenberg PB and
Williams RS, et al. Exercise stimulates PGC-1a transcription in skeletal
69 Teyssier C, Ma H, Emter R, Kralli A and Stallcup MR. Activation of
muscle through activation of the p38 MAPK pathway. J Biol Chem 2005,
nuclear receptor coactivator PGC-1a by arginine methylation. Genes Dev
52 Wu H, Kanatous SB, Thurmond FA, Gallardo T, Isotani E, Bassel-Duby
70 Rytinki MM and Palvimo JJ. SUMOylation attenuates the function of
R and Williams RS. Regulation of mitochondrial biogenesis in skeletal
PGC-1a. J Biol Chem 2009, 284: 26184 – 26193.
muscle by CaMK. Science 2002, 296: 349 – 352.
71 Canto´ C, Gerhart-Hines Z, Feige JN, Lagouge M, Noriega L, Milne JC
53 Zong H, Ren JM, Young LH, Pypaert M, Mu J, Birnbaum MJ and
Shulman GI. AMP kinase is required for mitochondrial biogenesis in skel-
modulating NADþ metabolism and SIRT1 activity. Nature 2009, 458:
etal muscle in response to chronic energy deprivation. Proc Natl Acad Sci
72 Kelly TJ, Lerin C, Haas W, Gygi SP and Puigserver P. GCN5-mediated
54 Olesen J, Kiilerich K and Pilegaard H. PGC-1a-mediated adaptations in
transcriptional control of the metabolic coactivator PGC-1b through lysine
skeletal muscle. Pflugers Arch 2010, 460: 153 – 162.
acetylation. J Biol Chem 2009, 284: 19945 – 19952.
55 Mortensen OH, Frandsen L, Schjerling P, Nishimura E and Grunnet N.
73 Patti ME, Butte AJ, Crunkhorn S, Cusi K, Berria R, Kashyap S and
PGC-1a and PGC-1b have both similar and distinct effects on myofiber
Miyazaki Y, et al. Coordinated reduction of genes of oxidative metabolism
switching toward an oxidative phenotype. Am J Physiol Endocrinol Metab
in humans with insulin resistance and diabetes: potential role of PGC1 and
NRF1. Proc Natl Acad Sci USA 2003, 100: 8466 – 8471.
56 Rodrı´guez-Calvo R, Jove´ M, Coll T, Camins A, Sa´nchez RM, Alegret M and
74 Semple RK, Crowley VC, Sewter CP, Laudes M, Christodoulides C,
Merlos M, et al. PGC-1b down-regulation is associated with reduced ERRa
Considine RV and Vidal-Puig A, et al. Expression of the thermogenic
activity and MCAD expression in skeletal muscle of senescence-accelerated
nuclear hormone receptor coactivator PGC-1a is reduced in the adipose
mice. J Gerontol A Biol Sci Med Sci 2006, 61: 773–780.
tissue of morbidly obese subjects. Int J Obes Relat Metab Disord 2004,
57 Koo SH, Satoh H, Herzig S, Lee CH, Hedrick S, Kulkarni R and Evans
RM, et al. PGC-1 promotes insulin resistance in liver through
75 Wilson-Fritch L, Nicoloro S, Chouinard M, Lazar MA, Chui PC, Leszyk J
PPAR-a-dependent induction of TRB-3. Nat Med 2004, 10: 530 – 534.
and Straubhaar J, et al. Mitochondrial remodeling in adipose tissue associ-
58 Wolfrum C and Stoffel M. Coactivation of Foxa2 through PGC-1b pro-
ated with obesity and treatment with rosiglitazone. J Clin Invest 2004,
motes liver fatty acid oxidation and triglyceride/VLDL secretion. Cell
76 Puigserver P and Spiegelman BM. Peroxisome proliferator-activated
59 Nagai Y, Yonemitsu S, Erion DM, Iwasaki T, Stark R, Weismann D and
receptor-g coactivator 1a (PGC-1a): transcriptional coactivator and meta-
Dong J, et al. The role of peroxisome proliferator-activated receptor
bolic regulator. Endocr Rev 2003, 24: 78 – 90.
gamma coactivator-1 beta in the pathogenesis of fructose-induced insulin
77 Choi CS, Befroy DE, Codella R, Kim S, Reznick RM, Hwang YJ and Liu
resistance. Cell Metab 2009, 9: 252 – 264.
ZX, et al. Paradoxical effects of increased expression of PGC-1a on
60 De Souza CT, Gasparetti AL, Pereira-da-Silva M, Arau´jo EP, Carvalheira
muscle mitochondrial function and insulin-stimulated muscle glucose
JB, Saad MJ and Boschero AC, et al. Peroxisome proliferator-activated
metabolism. Proc Natl Acad Sci USA 2008, 105: 19926 – 19931.
receptor gamma coactivator-1-dependent uncoupling protein-2 expression
78 Ek J, Andersen G, Urhammer SA, Gaede PH, Drivsholm T, Borch
in pancreatic islets of rats: a novel pathway for neural control of insulin
Johnsen K and Hansen T, et al. Mutation analysis of peroxisome
secretion. Diabetologia 2003, 46: 1522 – 1531.
proliferator-activated receptor-gamma coactivator-1 (PGC-1) and relation-
61 Zhang P, Liu C, Zhang C, Zhang Y, Shen P, Zhang J and Zhang CY. Free
ships of identified amino acid polymorphisms to type II diabetes mellitus.
fatty acids increase PGC-1a expression in isolated rat islets. FEBS Lett
Diabetologia 2001, 44: 2220 – 2226.
79 Mistry NF and Cresci S. PPAR transcriptional activator complex poly-
62 Kitamura T, Nakae J, Kitamura Y, Kido Y, Biggs WH, III, Wright CV and
morphisms and the promise of individualized therapy for heart failure.
White MF, et al. The forkhead transcription factor Foxo1 links insulin
Heart Fail Rev 2010, 15: 197 – 207.
PGC-1 coactivators in the control of energy metabolism
80 Zhang Y, Liu C, Zhu L, Jiang X, Chen X, Qi X and Liang X, et al.
86 Feilchenfeldt J, Bru¨ndler MA, Soravia C, To¨tsch M and Meier CA.
PGC-1a inhibits oleic acid induced proliferation and migration of rat vas-
Peroxisome proliferator-activated receptors (PPARs) and associated tran-
cular smooth muscle cells. PLoS One 2007, 2: e1137.
scription factors in colon cancer: reduced expression of PPARg-coactivator
81 Zhu L, Sun G, Zhang H, Zhang Y, Chen X, Jiang X and Jiang X, et al.
1 (PGC-1). Cancer Lett 2004, 203: 25 – 33.
PGC-1a is a key regulator of glucose-induced proliferation and migration
87 Ba Y, Zhang CN, Zhang Y and Zhang CY. Down-regulation of PGC-1a
in vascular smooth muscle cells. PLoS One 2009, 4: e4182.
expression in human hepatocellular carcinoma. Zhonghua Zhong Liu Za
82 Valle I, Alvarez-Barrientos A, Arza E, Lamas S and Monsalve M.
PGC-1a regulates the mitochondrial antioxidant defense system in vascu-
88 Zhang Y, Ba Y, Liu C, Sun G, Ding L, Gao S and Hao J, et al. PGC-1a
lar endothelial cells. Cardiovasc Res 2005, 66: 562 – 573.
induces apoptosis in human epithelial ovarian cancer cells through a
83 Taherzadeh-Fard E, Saft C, Andrich J, Wieczorek S and Arning L. PGC-1a as
PPARg-dependent pathway. Cell Res 2007, 17: 363 – 373.
modifier of onset age in Huntington disease. Mol Neurodegener 2009, 4: 10.
89 Lee HJ, Su Y, Yin PH, Lee HC and Chi CW. PPARg/PGC-1a pathway in
84 Handschin C, Kobayashi YM, Chin S, Seale P, Campbell KP and
E-cadherin expression and motility of HepG2 cells. Anticancer Res 2009,
Spiegelman BM. PGC-1a regulates the neuromuscular junction program and
ameliorates Duchenne muscular dystrophy. Genes Dev 2007, 21: 770– 783.
90 Zhang Y, Huypens P, Adamson AW, Chang JS, Henagan TM, Boudreau
85 Watkins G, Douglas-Jones A, Mansel RE and Jiang WG. The localization
A and Lenard NR, et al. Alternative mRNA splicing produces a novel bio-
and reduction of nuclear staining of PPARg and PGC-1 in human breast
logically active short isoform of PGC-1a. J Biol Chem 2009, 284:
cancer. Oncol Rep 2004, 12: 483 – 488.
Acta Biochim Biophys Sin (2011) | Page 10
Capítulo 2 MARCO CONCEPTUAL DEL ENVEJECIMIENTO EXITOSO, DIGNO, ACTIVO, PRODUCTIVO Y SALUDABLE Tomás Engler “Todavía tengo casi todos mis dientes casi todos mis cabellos y poquísimas canas puedo hacer y deshacer el amor trepar una escalera de dos en dos y correr cuarenta metros detrás del ómnibus o sea que no debería sentirme viejo pero el grave problema es que antes no
CONSTITUCIONALISMO, MINORÍAS Y DERECHOS1. 1. Introducción E n los últimos años hemos asistido a la inserción, dentro de los debates sobre el contenido y papel de las Constituciones, del tema del multi-culturalismo. No se trata del clásico enfoque sobre la protección cons-titucional que debía darse a las minorías, sino de una posición que ha ve-nido a poner en crisis varios d
 Acta Biochimica et Biophysica Sinica Advance Access published February 16, 2011
Acta Biochimica et Biophysica Sinica Advance Access published February 16, 2011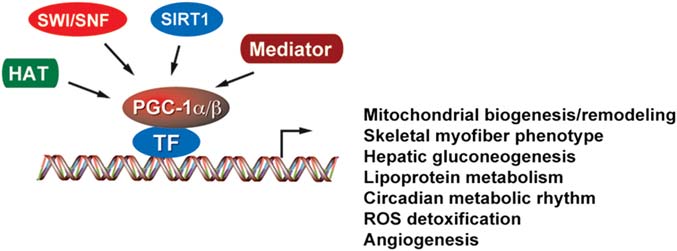 PGC-1 coactivators in the control of energy metabolism
domain is a regulatory region that roughly spans 200
energy demand, including the BAT, heart, skeletal muscle,
amino acids. Toward the C terminus, PGC-1a recruits the
kidney, and brain – In fact, when ectopically
thyroid receptor-associated protein/vitamin D receptor-
expressed in fat or muscle cells, PGC-1a strongly stimu-
interacting protein/mediator complex that facilitates direct
lates the program of nuclear and mitochondrial-encoded
interaction with the transcription initiation machinery [].
PGC-1 coactivators in the control of energy metabolism
domain is a regulatory region that roughly spans 200
energy demand, including the BAT, heart, skeletal muscle,
amino acids. Toward the C terminus, PGC-1a recruits the
kidney, and brain – In fact, when ectopically
thyroid receptor-associated protein/vitamin D receptor-
expressed in fat or muscle cells, PGC-1a strongly stimu-
interacting protein/mediator complex that facilitates direct
lates the program of nuclear and mitochondrial-encoded
interaction with the transcription initiation machinery [].