Levitra has a minimal amount of contraindications which has increased its popularity viagra uk You can buy quality certified medications from us at an affordable price.
Ppharmp503
C Basic & Clinical Pharmacology & Toxicology 2005, 97, 61–73. Printed in Denmark . All rights reserved MiniReview How Palatable Food Disrupts Appetite Regulation Charlotte Erlanson-Albertsson
Section of Endocrinology and Metabolism, Department of Experimental Medicine, University of Lund, Lund, Sweden
(Received September 15, 2004; Accepted March 7, 2005)
Abstract: Appetite regulation is part of a feedback system that controls the energy balance, involving a complex interplayof hunger and satiety signals, produced in the hypothalamus as well as in peripheral organs. Hunger signals may begenerated in peripheral organs (e.g. ghrelin) but most of them are expressed in the hypothalamus (neuropeptide Y, orexins,agouti-related peptide, melanin concentrating hormone, endogenous opiates and dopamine) and are expressed duringsituations of energy deficiency. Some satiety signals, such as cholecystokinin, glucagon-like peptide 1, peptide YY andenterostatin are released from the digestive tract in response to food intake. Others, such as leptin and insulin, aremobilized in response to perturbations in the nutritional state. Still others are generated in neurones of the hypothalamus(a-melanocyte-stimulating hormone and serotonin). Satiety signals act by inhibiting the expression of hunger signals and/or by blunting their effect. Palatable food, i.e. food rich in fat and sugar, up-regulates the expression of hunger signalsand satiety signals, at the same time blunting the response to satiety signals and activating the reward system. Hence,palatable food offsets normal appetite regulation, which may explain the increasing problem of obesity worldwide.
Today obesity is the greatest threat to public health in the
pears to be a highly regulated process, considering the irregu-
developed world. It is a silent killer in many disorders, such
larity in the number and composition of meals from one day
as heart disease, hypertension and diabetes that are the
to the next, and the energy balance and stability in body
consequences of obesity (Taubes 1998). Genetic factors are
weight over longer periods of time. On the other hand obesity
certainly important for the development of obesity. This is
in the world is increasing, suggesting that there is a funda-
true for a large number of mutations, for instance those
mental weakness in the regulation of appetite and in the con-
occurring in the melanocortin receptor gene (Yeo et al.
trol of energy homeostasis. The aim of this article is to review
2003). Hence, it has been found that mutations in the mela-
the mechanisms underlying appetite regulation and the in-
nocortin receptor gene result in a distinct obesity syndrome
fluence of highly palatable food on this process. This review
that is inherited in a co-dominant manner (Farooqi et al.
draws from previous overviews on the control of hedonic eat-
2003). Various other mutations are associated with obesity
ing (Saper et al. 2002; Cummings & Schwartz 2003; Zig-
as illustrated by leptin gene deficiency (Montague et al.
man & Elmquist 2003; Gale et al. 2004).
1997) or leptin receptor gene deficiency (Clement et al. 1998). Although such mutations verify the importance of
General aspects of hunger and satiety
genetic factors, they do not explain obesity world-wide(Froguel et al. 2000).
The regulation of food intake is based on an intricate feed-
The most important cause of obesity is probably over-eat-
back system, which is influenced not only by the availability
ing (coupled with inactivity). Over-eating is stimulated by the
of nutrients but also by various environmental and psycho-
ready availability of food rich in fat and sugar. Appetite regu-
logical factors. It is a complex system, as has been described
lation and the understanding of the rewarding effect of food,
in a number of recent reviews (Schwartz et al. 2000; Blund-
in particular energy-rich food (fat and sugar), has become of
ell & Gillett 2001). Basically however, food intake is con-
great scientific interest. Much knowledge has been provided
trolled by hunger and satiety signals. The signals are gener-
by techniques of molecular biology, which have identified a
ated in peripheral organs, such as the digestive tract and
large number of genes and gene products responsible for sen-
adipose tissue, and in the brain itself. Certain neurones in
sations of hunger and satiety. On one hand food intake ap-
the hypothalamus are the targets of these signals. The fairlyrecent identification of hunger- and satiety peptides as well
Author for correspondence: Charlotte Erlanson-Albertsson, Divi-
as their receptors has renewed the interest in appetite regu-
sion of Diabetes, Metabolism and Endocrinology, Department of
lation with special focus on the identification of tools that
Experimental Medicine, University of Lund, Biomedical Center(BMC), F13, SE-221 84 Lund, Sweden (fax π46 46 2224022, e-mail
might be useful in the pharmacological treatment of eating
Charlotte.Erlanson-Albertsson/med.lu.se).
disorders, from anorexia to over-eating and obesity. MiniReview
(Tschop et al. 2000; Dornonville de la Cour et al. 2001;
Hunger signals and the drive to eat
Wren et al. 2001 & 2002). It is produced in the stomach by
The identification of appetite regulating signals started with
endocrine cells (A-like cells) in the oxyntic glands and se-
the realization that satiety peptides signals are of paramount
creted into the blood stream (Dornonville de la Cour et al.
importance in appetite control (to terminate food intake). It
2001). The pre-prandial rise in ghrelin is not proportional
has since become clear that the sensation of hunger, the drive
to the ingested caloric load during the previous meal
to seek food, is the central event in appetite regulation, satiety
(Callahan et al. 2004). Ghrelin receptors occur in the arcu-
signals inhibiting the effect of hunger signals. Quite a few
ate nucleus, where they activate neuropeptide Y neurones
hunger peptides have been identified, among these neuropep-
(Tschop et al. 2002). It has also been shown that ghrelin
tide Y, ghrelin, the orexins and agouti-related peptide, and
releases neuropeptide Y from hypothalamic explants in vi-tro, suggesting that it may be a mediator of the ghrelin ef-
Neuropeptide Y is a 36 amino acid peptide with sequence
fect (Wren et al. 2002). A role of endogenous ghrelin in
homologies with pancreatic polypeptide and peptide YY. It
appetite regulation is suggested by an increased ghrelin ex-
is abundantly expressed in both the central and peripheral
pression in the stomach following fasting and a decrease
nervous system (Allen et al. 1983), a great density of neuro-
in circulating ghrelin after intake of a regular chow meal
peptide Y neurones being observed in the arcuate nucleus,
(Dornonville de la Cour et al. 2001; Kim et al. 2003). The
projecting to the paraventricular nucleus. Injection of
suppression of plasma levels of ghrelin following a meal
neuropeptide Y into the paraventricular hypothalamus
may be an effect of nutrients in blood. The most striking
stimulates food intake in the rat, carbohydrates in particu-
effect of long-term ghrelin treatment is the increased body
lar (Stanley & Leibowitz 1985). Upon intracerebroventricul-
weight and accumulation of fat (Tschop et al. 2000). That
ar injection neuropeptide Y suppresses energy expenditure
ghrelin may be involved in the defense against human obes-
from brown adipose tissue (Billington et al. 1994). A
ity is suggested by the observation of decreased blood levels
physiological role of neuropeptide Y is suggested by the fact
of ghrelin in obese subjects, as demonstrated in obese Pima
that it is released from the hypothalamus in situations of
Indians who were found to have 33% lower fasting plasma
fasting and in situations of increased energy demands, such
ghrelin than lean subjects (Tschop et al. 2001). Ghrelin has
as exercise and lactation. Long-term infusion of neuropep-
been shown to increase food intake in humans (Wren et al.
tide Y into the lateral ventricle of mice produces hyper-
2001), demonstrated after a 270 min. infusion of ghrelin,
phagia, insulin resistance, and obesity (Raposinho et al.
during which a standard free-choice buffet was served.
2001). Raised neuropeptide Y mRNA levels in specific hy-
Ghrelin expression in the mouse stomach is decreased in
pothalamic nuclei, including the arcuate nucleus and the
response to a high-fat diet (Moesgaard et al. 2004), an effect
paraventricular nucleus are found in animals with hyper-
that may serve to restrain the hyperphagia induced by high-
phagia and obesity (Gao et al. 2002). Upon fasting the
fat diets. The effect of sucrose on ghrelin expression is not
neuropeptide Y knockout mouse has a lower food intake
than wild-type mice, nut normal body weight (Bannon etThe orexins are two homologous peptides A and B, iden-
al. 2000), supporting a role for neuropeptide Y in mediating
tified as ligands for two orphan receptors, belonging to the
energy intake and energy homeostasis. Both leptin and in-
G-protein coupled receptor family (Sakurai et al. 1998).
sulin inhibit the expression of neuropeptide Y and its recep-
The two peptides derive from the same 130 amino acid pre-
tor (Schwartz et al. 1991; Erickson et al. 1996), which can
cursor, prepro-orexin, that is cleaved into orexin A and B.
be interpreted to mean that hunger elicited by neuropeptide
They are produced in neurones of the lateral and posterior
Y is controlled by leptin and insulin.
hypothalamus, which project widely in the brain. Intracere-
High-fat diets have been found to either reduce the ex-
broventricular injection of orexin A or B promptly stimu-
pression of neuropeptide Y in neurones of the arcuate nu-
lates food intake (Sakurai et al. 1998). There is a close inter-
cleus (Giraudo et al. 1994; Lin et al. 2000) or to stimulate
action between neuropeptide Y, orexin and leptin, the ex-
neuropeptide Y expression (Huang et al. 2004). The reason
pression of orexins being increased by neuropeptide Y and
for the conflicting data is not clear, but the type of fat in-
suppressed by leptin (Niimi et al. 2001). Orexin neurones
gested is probably important, saturated fat producing an
are activated by fasting (Diano et al. 2003).
upregulation of neuropeptide Y expression in contrast to
Orexin gene expression is stimulated by high plasma tri-
polyunsaturated fat (Huang et al. 2004). A sucrose-contain-
glyceride levels, obtained either through high-fat consump-
ing diet has been found to stimulate the expression of hypo-
tion or through Intralipid infusion (Wortley et al. 2003).
thalamic neuropeptide Y, and to cause hyperglycaemia,
The effect of sucrose on orexin expression is not known.
hyperinsulinaemia and an increased body weight (Kaga etAgouti-related peptide is a neuropeptide, synthesized and
al. 2001). Since neuropeptide Y stimulates food intake an
secreted by neuropeptide Y neurones in the arcuate nucleus
upregulation of hypothalamic neuropeptide Y by saturated
(Korner et al. 2001). Agouti-related peptide stimulates appe-
fat or by sucrose may be one reason for the hyperphagia
tite and causes obesity as first described in the yellow agouti
mouse having an overexpression of agouti protein (Klebig etGhrelin is a meal initiator, circulating levels being elev-
al. 1995). Agouti-related peptide interacts with the mela-
ated during fasting and suppressed following a meal
nocortin receptor, acting as a competitive antagonist to a-
MiniReview
PALATABLE FOOD DISRUPTS APPETITE REGULATION
melanocyte-stimulating hormone (a-MSH), which is an in-
food intake in experimental animals, some of them, like
hibitor of food intake. Administration of agouti-related pep-
cholecystokinin, glucagon-like peptide 1 and peptide YY,
tide to hypothalamic explants stimulated the release of
have been shown to reduce food intake also in man (Gutz-
neuropeptide Y, suggesting that orexigenic behaviour can be
enforced via a positive feedback loop (Dhillo et al. 2002). Cholecystokinin was the first peptide hormone to be as-
Mice on a high-fat diet have higher agouti-related peptide
sociated with meal-evoked satiety (Smith & Gibbs 1975).
mRNA expression in the bed nucleus of the stria terminalis
Cholecystokinin is released in response to nutrients in the
and in the ventral part of the lateral septal nucleus than
duodenum and acts by stimulating vagal afferents, which
mice on a low-fat diet suggesting that agouti-related peptide
carry cholecystokinin-1 receptors (Moran et al. 1997). Cho-
may be involved in fat-evoked hyperphagia (Huang et al.
lecystokinin is thought to act as a neurotransmittor or
2003). The effect of sucrose on agouti-related peptide ex-
neuromodulator in various parts of the brain, for instance
in the nucleus tractus solitarius and in the medial basal
Melanin-concentrating hormone is a proteolytic cleavage
hypothalamus. Following intracerebroventricular injection
product of the melanin-concentrating hormone preprohor-
of cholecystokinin food intake is reduced, whereas injection
mone of 165 residues, giving rise also to two other peptides,
of a cholecystokinin-1-receptor antagonist stimulates food
neuropeptide E-I and neuropeptide G-E (Shi 2004). Mela-
intake (Corp et al. 1997). One important effect of cholecys-
nin-concentrating hormone-expressing neurones in the
tokinin is to suppress the neuropeptide Y expression in the
brain have a restricted localization to the lateral hypothala-
dorsomedial hypothalamus (Bi et al. 2001). Rats lacking the
mus area and zona incerta (Zamir et al. 1986; Bittencourt
cholecystokinin-1 receptor, the Otsuka Long-Evans Toku-
et al. 1992) with projections to the nucleus of the solitary
shima Fatty (OLETFA) rats have a five-fold elevation of
tract. Cumulative evidence suggests that melanin-concen-
neuropeptide Y expression in this region, which could ex-
trating hormone plays an important role in maintaining en-
plain their hyperphagia and obesity. Thus, cholecystokinin
ergy homeostasis by regulating food intake and energy ex-
seems to be important within the brain to regulate hunger
penditure (Shi 2004). Intracerebroventricular injection of
melanin-concentrating hormone stimulates food intake in
There is no data on the expression of cholecystokinin in
rats (Rossi et al. 1997; Shearman et al. 2003) and long-
the upper small intestine after high-fat or sucrose diets. The
term infusion of melanin-concentrating hormone induces
satiety response to circulating cholecystokinin is blunted in
hyperphagia and body weight gain (Ito et al. 2003). The
rats maintained on a high-fat diet compared to rats main-
expression of melanin-concentrating hormone is up-regu-
tained on a low-fat diet (Covasa et al. 2000). Also, inhi-
lated by food deprivation (Qu et al. 1996; Herve &
bition of gastric emptying by cholecystokinin is markedly
Fellmann 1997). It is also upregulated in the leptin-deficient
attenuated in rats maintained on a high fat diet. The attenu-
ob/ob mouse, suggesting that leptin controls the expression
ation of cholecystokinin-induced inhibition of food intake
of melanin-concentrating hormone (Segal-Lieberman et al.
and gastric emptying is thought to reflect a reduced vagal
2003). In addition to being present in the hypothalamus,
cholecystokinin responsiveness, measured as a decreased
melanin-concentrating hormone is also present in periph-
Fos expression in nuclei of the solitary tract and area post-
eral tissues and in blood (Sun et al. 2004). There is a posi-
rema, where the vagal sensory fibers terminate (Covasa et
tive correlation between serum melanin-concentrating hor-
al. 2000). There is also a blunted satiety response to chole-
mone levels and body mass index and fat mass (Gavrila
cystokinin during hyperglycemia (Lam et al. 1998). et al. 2005), in line with the view that circulating melanin-
Glucagon-like peptide 1 is a gut hormone derived from
concentrating hormone acts as a hunger signal.
the processing of proglucagon in intestinal L cells. In ad-dition to stimulating insulin release, glucagon-like peptide 1also reduces food intake after intracerebroventricular ad-
Satiety signals and the termination of feeding
ministration (Turton et al. 1996). The glucagon-like peptide-
The sensation of satiety (meal termination) reflects the sup-
receptor knock-out mouse did not display obesity or in-
pression of hunger signals and/or the mobilization of satiety
creased food intake, possibly because other satiety systems
signals in response to a meal. Many of these latter signals
were activated (Gallwitz & Schmidt 1997). Glucagon-like
derive from the pancreas and gastrointestinal tract, trans-
peptide 1 is of particular interest because it inhibits food
mitting information from the periphery (via the vagus or
intake also in man (Naslund et al. 1999). The mechanism
via the circulation) to the brain. Other circulating satiety
behind the glucagon-like peptide 1-induced inhibition of
signals (nutrient metabolites) are generated in the liver, en-
food intake is probably a combined effect of gastric dila-
tering the brain from the blood (e.g.glucose and ketone
tation and an increase in the level of circulating serotonin,
associated with satiety (Owji et al. 2002). In dogs fed a high-fat diet the fasting plasma glucagon-like peptide 1 concen-
Meal-initiated satiety signals from the digestive tract: chole-
tration was 2.5 times higher than in controls (van Citters
cystokinin, glucagon-like peptide 1 and peptide YY.et al. 2002). Additionally, expression of the glucagon-like
Gastrointestinal satiety peptides are released from the intes-
peptide 1 receptor in the whole pancreas was increased 2.3
tine in response to food. While all of these peptides reduce
times in the fat-fed animals. This suggests that glucagon-
MiniReview
like peptide 1 may provide a defence against hyperphagia
Leptin has emerged as a major suppressor of appetite and
induced by high-fat food. The effect of sucrose on gluca-
is therefore viewed as an anti-obesity hormone (Fried-
gon-like peptide 1 expression is not known.
man & Halaas 1998; Leibel 2002; Loos & Bouchard 2003). Peptide YY is released from the intestinal L cells to re-
This view is based on the finding that total deficiency of
duce food intake in rodents as well as in man (Batterham
leptin or its receptor leads to hyperphagia and obesity in
et al. 2002). It acts to inhibit the electrical activity of neuro-
mice and man (Campfield et al. 1995; Montague et al.
peptide Y neurones in the arcuate nucleus (Batterham et al.
1997). Leptin is a 16-kDa protein produced in adipose
2002). It has been suggested that peptide YY serves as a
tissue and secreted into the blood. Leptin has its own recep-
circulating factor mediating satiety following ingestion of a
tor with signalling properties similar to the interleukin-6
meal (Batterham et al. 2002). Peptide YY is active as a sat-
receptor, localized to the arcuate nucleus (Tartaglia et al.
iety signal with both high-fat and low-fat diets (Challis et al.
1995). Leptin is thought to pass the blood-brain barrier to
2004) and hence may provide a defence against fat-induced
reach its receptor, reducing food intake and increasing ther-
hyperphagia. Whether peptide YY acts as a satiety signal
mogenesis (Friedman & Halaas 1998). The main role of lep-
with sucrose-enriched food is unknown. Endogenous post-
tin is to control the expression and activity of various other
prandial levels of peptide YY were significantly lower in
appetite controlling peptides. One target for leptin is the
obese subjects compared to a lean group. Peptide YY in-
arcuate nucleus, which harbours neuropeptide Y neurones
fusion also caused a significant decrease in the cumulative
that carry receptors for both leptin and insulin (Erickson
24 hr caloric intake in both obese and lean subjects, sug-
et al. 1996). Fasting or weight loss lead to low blood leptin
gesting that obese subjects were sensitive to the peptide YY
levels, which in turn cause the hypothalamic neuropeptide
action (Batterham et al. 2003).
Y expression to rise, thereby stimulating food intake (Blun-
Adiposity related satiety signals: insulin, leptin and pro-opiomelanocortin (POMC)-derived peptides. Inter-meal and
High-fat diet causes increased leptin expression and an
long-term signalling constitutes a complementary regula-
increased body weight (20%) compared with standard diet
tory system that probably acts synergistically to short-term
as illustrated in rodents (Moraes et al. 2003). This suggests
that leptin is involved in the feedback control of fat intake.
The ability of insulin to reduce food intake at first ap-
High-fat diets result in an inability to respond to leptin
peared a paradox, insulin being a hormone that promotes
(Moraes et al. 2003). This phenomenon will be discussed
storage of energy in liver and adipose tissue. The paradox
below under the name of leptin resistance. Leptin has also
was resolved, when it was found that insulin induced satiety
been found to inhibit the response to sweet taste (Kawai et
by acting on receptors in the brain, whereas the anabolic
al. 2000), suggesting that the intake of sweet food might be
actions of insulin occurred peripherally (Woods et al. 2000).
regulated by leptin. A high-sucrose meal was found to in-
There is evidence that insulin passes the blood-brain barrier
crease adipose tissue leptin mRNA levels by at least 5 times
intact and in a regulated receptor-mediated fashion (Woods
within 3 hr, suggesting a feedback control of leptin to regu-
et al. 2003). When blood insulin is raised following a meal,
late sucrose intake (Polson & Thompson 2003). The en-
the passage of insulin into the brain is thought to reflect
hanced preference for sweet substances found in the db/db
this increase (Baskin et al. 1999). After fasting the penetra-
mice lacking the leptin receptor could hence be explained
tion of insulin into the brain is greatly reduced with the
by a defect leptin suppression of the reward induced by the
consequence that a larger meal can be eaten before insulin
sweet taste (Shigemura et al. 2004). In a comparison be-
evokes satiety (Strubbe et al. 1988).
tween dietary sucrose and coconut fat (rich in saturated
High-fat diets induce insulin resistance, which from the
fatty acids) in ob/ob mice, lacking leptin, it was found that
point of view of appetite regulation means suppressed satiety
the body weight gain was greater in sucrose-fed ob/ob mice
(Kim et al. 2004). That insulin indeed inhibits the intake of
than in fat-fed mice (Flatt et al. 1990). Further experiments
high-fat food is clear from the use of non-peptide insulin mi-
are needed to establish a role of leptin in promoting satiety
metics, which when given intracerebroventricular were
shown to reduce over-eating and prevent obesity in rats main-
Pro-opiomelanocorticotropin is a prohormone produced
tained on high-fat diet (Air et al. 2002). Intracerebroventricu-
by specific neurones in the arcuate nucleus that seem to op-
lar injection of insulin has been shown to reduce sucrose in-
erate under leptin control. Approximately 40% of the pro-
take in rats, that have been stimulated to eat sucrose by treat-
opiomelanocorticotropin neurones carry leptin receptors
ment with the opiate k receptor agonist U50, 488 (Sipols et
(Cheung et al. 1997). The pro-opiomelanocorticotropin
al. 2002). However, sucrose-rich diets are known to cause the
neurones generate a-melanocyte-stimulating hormone, which
classical signs of insulin resistance, with increased food in-
is released from terminals in the paraventricular nucleus
take, adiposity, hyperinsulinaemia and hypertriglyceridaem-
and lateral hypothalamus (Tritos & Maratos-Flier 1999).
ia (Davidoff et al. 2004), indicating that the defence against
a-melanocyte-stimulating hormone suppresses food intake,
over-eating afforded by insulin is overridden at some point.
acting through the melanocortin-4 receptor in neurones of
Also the passage of insulin through the blood brain barrier is
the arcuate nucleus. The leptin-induced activation of the
decreased after a high-fat meal compared to a low-fat meal,
melanocortin receptor seems to be critical for the regulation
hence leading to a reduced satiety (Gerozissis et al. 1997).
of food intake, since interruption at any point in this chain
MiniReview
PALATABLE FOOD DISRUPTS APPETITE REGULATION
of events causes overeating (Yeo et al. 1998). Leptin stimu-
Palatable food and appetite regulation
lates the pro-opiomelanocorticotropin neurons, while inhib-iting the neuropeptide Y neurones (Williams et al. 1999).
As stated above, palatable food, i.e. food rich in fat and
Hence, the arcuate nucleus of the hypothalamus controls
sugar, will increase food intake, i.e. the activity and expres-
food intake, acting through neuropeptide Y neurones to
sion of signals controlling appetite will be balanced in fa-
stimulate and through pro-opiomelanocorticotropin neur-
vour of prolonged eating (table 2). Palatable food activates
ones (a-melanocyte-stimulating hormone) to inhibit food
the reward system, thereby affecting ingestive behaviour.
intake. Patients with a defect of the pro-opiomelanocortico-
Typical of ‘‘reward eating’’ is that the driving force is grati-
tropin gene product due to mutations were found to display
fication rather than energy deficit (Pelchat 2002). From an
red hair, early-onset obesity, and congenital hypocortisolism
evolutionary point of view it makes sense that food rich in
(Krude et al. 2003; Farooqi & O’Rahilly 2004). Also, in-
fat and sugar is attractive, because such food can be rapidly
herited abnormalities in the synthesis and processing of
converted into energy (Nesse & Berridge 1997). Long-term
pro-opiomelanocorticotropin and defects in the action of
overconsumption of palatable food has been compared to
pro-opiomelanocorticotropin-derived peptides could help
drug addiction (Berridge 1996; Gosnell 2000). The behav-
to explain obesity (Coll et al. 2004).
iour induced by stimulating the reward system is to ‘‘come
With a high-fat diet pro-opiomelanocorticotropin-de-
back for more’’ (Kelley et al. 2002). Accordingly, free access
rived peptides appear to be important to enable the individ-
to palatable food may lead to over-eating, characterized by
ual to resist overeating. This conclusion is based on studies
prolongation of the meal because the normally induced sen-
of pro-opiomelanocorticotropin-deficient mice, which re-
sation of satiety is overridden. Another parallel between
spond to a high-fat diet with hyperphagia and obesity
palatable food and addictive drugs is the phenomenon of
(Challis et al. 2004). The importance of pro-opiomelanocor-
adaptation (Koob & Le Moal 1997). Addictive drugs are
ticotropin-derived peptides in appetite regulation is sup-
known to start a series of adaptations leading to a shift in
ported by studies of mice on a high-fat diet for 13 weeks;
homeostatic set points. Overconsumption of palatable food
they became obese concomitantly with a reduction of pro-
gradually shifts the set point for energy balance and body
opiomelanocorticotropin mRNA expression in the arcuate
nucleus (Huang et al. 2004). Pro-opiomelanocorticotropin
There are two main explanations for compulsive over-
mRNA in these animals was upregulated by a diet rich in
eating of palatable food. One is the activation of the reward
n-3 polyunsaturated fat, emphasising the fact that the type
system, represented by endogenous opioids, dopamine and
of dietary fat matters in the regulation of hypothalamic
serotonin. The other is the phenomenon of ‘‘resistance‘‘, i.e.
neuropeptide expression (Huang et al. 2004).
an impaired ability to respond to food intake with a signal-
During the processing of the pro-opiomelanocorticotrop-
in molecule, endorphins are also produced, which stimulaterather than inhibit appetite. Mice lacking b-endorphin butnot the other pro-opiomelanocorticotropin-derived pep-
tides were orexigenic, suggesting that endorphins are im-
Effects of diets rich in fat or sucrose on the expression of hunger
portant in normal appetite regulation and that the different
pro-opiomelanocorticotropin-derived peptides interact in a
complex manner in the regulation of energy homeostasis
Peptides involved in appetite control.
Hunger peptides are released in the circulation or upregulated
(hypothalamus) in response to food deprivation. Satiety peptides
are released into the circulation or upregulated (hypothalamus) in
NPY – neuropeptide Y, AgRP – agouti-related peptide, MCH –
melanin-concentrating hormones, MSH – melanocyte-stimulatinghormone, CCK – cholecystokinin, GLP – glucagon-like peptide 1,
↑ upregulated, ↓downregulated, – not investigated, * saturated fat. MiniReview
eners, while dynorphin and U 50 488, two k-agonists, did
Activation of the reward system by palatable food
not (Zhang & Kelley 1997 & 2002). Water intake was not
The reward system. In recent years the mechanisms underly-
affected, indicating that opioids are not involved in the con-
ing reward and motivation have been outlined (Kelley
trol of water intake. In man, the opiate receptor antagonist
2004). Nucleus accumbens is a brain region that appears to
naloxone was found to reduce the preference for palatable
play a crucial role in behaviour related to natural re-
food, without affecting subjective ratings of hunger and sat-
inforcers, such as ingestion of food and mating. The nucleus
iety (Drewnowski et al. 1992). It is to be noted that opiate
accumbens is also a site for the rewarding and reinforcing
antagonists decrease the attractiveness of food without af-
properties of addictive drugs. The nucleus accumbens re-
fecting its taste, i.e. the subjects are still able to sense the
ceives information from the brain stem in response to taste
sweet taste, but it does not elicit the expected reward (Yeo-
and gastrointestinal sensations from the food ingested
mans & Gray 1996; Kelley et al. 2002). Opioids stimulate
through a direct connection with the nucleus of the solitary
food intake because the sensation of hunger is more intense.
tract. There is also a connection between the nucleus ac-
This conclusion was drawn from experiments in which rats
cumbens and the lateral hypothalamus, being important for
had to press a bar to receive food (Glass et al. 1999). Upon
energy homeostasis. Although there are many neurotrans-
intravenous infusion of opioid agonists the number of bar
mitter systems within this region, studies on the rewarding
presses (i.e. the urgency) increased before the rats gave up
effects of food has focused on three signal systems, involv-
(Glass et al. 1999). In contrast naloxone reduced the ur-
ing endogenous opioids, dopamine and serotonin.
gency of the food-seeking behaviour. Together these obser-
The opioid system. Since the discovery of the endogenous
vations suggest that endogenous opioids are important to
opioid peptides and their receptors nearly 30 years ago
there has been a need to understand the role of these com-
Upon ingestion of palatable food the hypothalamic ex-
pounds in the brain (Pert & Snyder 1973; Lord et al. 1977).
pression of opioid peptides and opioid receptors is in-
Initially, interest was focused on pain and pain relief but
creased, supporting the involvement of the opioid system in
lately has included also reward, addiction and ingestive be-
palatable food consumption. High-fat and sucrose diets
haviour. In mammals the endogenous opioid peptides are
were shown to increase the expression of prodynorphin/dy-
derived from the proteins pro-opiomelanocortin (POMC),
norphin in the arcuate nucleus and paraventricular nucleus
proenkephalin, prodynorphin and pronociceptin/orphanin
(Kim et al. 1996; Welch et al. 1996). At the same time intra-
FQ. These precursors produce the opioid peptides b-endor-
cerebroventricular injection of opioids stimulated the intake
phins, enkephalins, dynorphins and nociceptin. b-Endorphin
of sucrose or sweeteners (Zhang & Kelley 2002) and fat
binds with equal affinity to m- and d-receptors, whereas dy-
(Ookuma et al. 1997). While opioids induce an increased
norphins bind to the k-receptor. The opiate receptors are
appetite for palatable food, the intake of palatable food is
found in various neuronal networks in the brain, for in-
decreased by opiate receptor antagonists. Enterostatin is a
stance in hypothalamic regions involved in the control of
peptide produced by proteolytic cleavage of pancreatic pro-
food intake and in the nucleus accumbens (reward centre).
colipase in the gut and has been found to inhibit fat intake
One important property of opiates is to reinforce the behav-
in an experimental rat model, provided the animals have
iour best described as ‘‘coming back for more’’ (Van Ree et
been adapted to fat feeding during 14 days (Erlanson-Al-
al. 2000). Systemic injection of morphine causes rats to
bertsson & York 1997). The mechanism of action for
overeat, an effect that can be prevented by various opiate
enterostatin occurs through an inhibition of the k and m
antagonists. When analyzing the type of food chosen under
opioid systems to reduce fat intake. The target protein for
the influence of opiates, it was found that morphine stimu-
enterostatin was however found to be the F1-ATPase b-sub-
lates the ingestion of food rich in sugar and fat (Yeomans
unit, enterostatin acting to inhibit ATP-production (Berger
et al. 1997). That it is the palatability of the food rather
et al. 2002 & 2004; Park et al. 2004). This suggests that
than its energy content that activates the opioid system is
an ATP-dependent intracellular mechanism is important for
clear from experiments comparing diets rich in sugar with
regulation of fat intake, opening new fields of investigation
diets rich in artificial sweeteners. Such studies show that the
sweet taste is important for opioid activation and not the
Dopamine. Another neurotransmittor that seems to be
energy content (Apfelbaum & Mandenoff 1981). It has also
involved in the reward response to food is dopamine. Several
been shown that the involvement of the opioid system dur-
different dopaminergic pathways are thought to modulate
ing intake of sweets is manifested as analgesia (Le Magnen
eating behaviour (Wang et al. 2001; Del Parigi et al. 2003). et al. 1980). Palatable food activates the opioid reward sys-
Five subtypes of dopamine receptors mediate the effects of
tem in the nucleus accumbens (Zhang et al. 2003). The nu-
dopamine, subtype 2 being associated with food intake and
cleus accumbens receives input from the medial prefrontal
reward (Blum et al. 2000). The dopaminergic circuitry is a
cortex, hippocampus and amygdala as well as from the
target for several appetite-regulating peptides, like galanin
brain stem, while output signals that promote eating target
(Rada et al. 1998), leptin (Szczypka et al. 2000) and chole-
the hypothalamus (Helm et al. 2003). In the rat microinjec-
cystokinin (Vaccarino 1994). Thus, microinjection of gala-
tion of various m-opiate agonists including morphine, en-
nin into the nucleus accumbens stimulates dopamine and
hanced sucrose intake as well as intake of artificial sweet-
suppresses acetylcholine release, i.e. signals for initiation of
MiniReview
PALATABLE FOOD DISRUPTS APPETITE REGULATION
feeding (Rada et al. 1998). Leptin was found to lower dop-
signals are down-regulated (Huang et al. 2003), thus lowering
amine release in the nucleus accumbens during feeding, sug-
the satiety response to a fat meal. Such changes in peptide ex-
gesting that leptin suppresses the feeding-induced reward
(Krugel et al. 2003). Dopamine is also involved in other
Other hunger signals like ghrelin are down-regulated in
aspects of eating behaviour, such as arousal and food seek-
response to intake of fat (Moesgaard et al. 2004). A high-
fat diet, on the other hand, upregulates several satiety sig-
Serotonin. A third neurotransmittor that may contribute
nals like leptin (Moraes et al. 2003), insulin, glucagon-like
to appetite regulation is serotonin. Several observations sug-
peptide 1 (van Citters et al. 2002) and enterostatin (Erlan-
gest that serotonin may be involved in the control of food in-
son-Albertsson & York 1997). Since a palatable food regime
take as a satiety signal (Lawton et al. 1995). Brain serotonin
leads to over-eating in spite of the change in appetite signals
levels are affected by many factors, including circulating
to restrict food intake, there must be either a blunted re-
levels of tryptophan and certain macronutrients (Halford &
sponse to satiety signals or food intake is stimulated by
Blundell 2000). In the hypothalamus, serotonin inhibits
other factors acting within the reward system. High serum
neuropeptide Y expression, thus depressing hunger (Half-
concentrations of leptin were noted in obese individuals
ord & Blundell 2000). Whether serotonin specifically regu-
(Maffei et al. 1995). There was also an inability of leptin to
lates carbohydrate intake (Wurtman & Wurtman 1995) and/
inhibit food intake in such individuals, a phenomenon
or fat intake (Blundell & Lawton 1995) is debated. Dietary
called ‘‘leptin resistance’’ (Frederich et al. 1995; Leibel
carbohydrates have been shown to raise brain serotonin turn-
2002). It was found that a high-fat diet caused a sustained
over as observed in depressed patients overeating carbo-
increase in circulating leptin in mice, and that the leptin
hydrates to increase well-being (Wurtman & Wurtman 1995).
levels reflected the amount of fat in the body (Frederich et
On the other hand treatment with the anorexic agent fenflu-
al. 1995). However, despite increased leptin levels, animals
ramine (Bray 2001), which is claimed to act by releasing sero-
on a high-fat diet became obese, suggesting that the high-
tonin as well as inhibiting serotonin reuptake, has been
fat diet changed the ‘set point’ for body weight/body fat, at
shown to specifically reduce fat intake (Lawton et al. 1995).
least in part by restraining the action of leptin (Frederich
Opioid antagonism and serotonergic stimulation, using
et al. 1995). Another explanation for the development of
naloxone and fluoxetin (a selective serotonin reuptake inhibi-
leptin resistance is an impaired ability of leptin to pass the
tor), has been shown to reduce intake of fat and sucrose in
blood-brain barrier (Banks et al. 1999; Banks & Farrell
animals (Hagan et al. 1997). That serotonin is important to
2003). The depressed passage of leptin in obese individuals
relieve stress was demonstrated in experiments where diets
is supposed to be a consequence of a high-fat diet. A third
rich in fat and carbohydrates were given to animals subjected
possible explanation for leptin resistance is inhibition of
to stress (Buwalda et al. 2001). A high-fat diet was found to
post-receptor signalling. Such an inhibitor of leptin signal-
reduce some of the behavioural responses to stressors more
ling has been identified, the SOCS-3, (suppressor-of-cyto-
effectively than a carbohydrate-based diet. It was also found
kine-signalling) (Bjorbaek et al. 1998), being upregulated by
that desensitization of central nervous 5-HT1a receptors oc-
high-fat diet and proposed to be a leptin resistance signal
curred in animals receiving a carbohydrate-based diet but
was absent in fat- fed animals (Buwalda et al. 2001). This ex-
The passage of insulin into the brain seems to be a key
periment supports a role for serotonin in relieving stress, but
event in insulin-induced satiety. There are specific regions in
also explains the urge for eating fat to relieve stress.
the brain, e.g. the hypothalamus and the hindbrain, whereinsulin penetration occurs (Banks & Kastin 1998). That pal-atable diet may reduce insulin penetration through the
Resistance mechanisms in appetite regulation
blood brain barrier was demonstrated in animals fed a
with palatable food
high-fat diet for several weeks (Burguera et al. 2000). This
In addition to activation of the reward system another ex-
could contribute to the development of obesity in individ-
planation for compulsive consumption of palatable food is
uals on a high-fat diet (Banks 2003). One might speculate
a blunted response to satiety signals. This may occur in dif-
that a high-fat diet causes a peripheral insulin resistance,
ferent ways: 1) an increased expression of hunger signals or
which shuffles glucose to the brain. Gradually the blood-
their receptors 2) a reduced expression of satiety signals and
brain barrier becomes resistant to the penetration of insulin
their receptors and 3) a default receptor targeting and/or
and the satiating effect of insulin is lost as a result.
post-receptor signalling in response to palatable food, in
A blunted satiety response to cholecystokinin seems to
develop in response to intake of food rich in fat (Covasa et
An altered expression upon long-term exposure to palat-
al. 2000) and sucrose (Lam et al. 1998). The mechanism
able food has been described for several of the appetite regu-
behind the blunted response is not known.
lating peptides listed in table 1. Some of the hunger peptideslike neuropeptide Y (Huang et al. 2004), the orexins (Wortley
Does palatable food cause addiction? et al. 2003) and agouti-related peptide (Huang et al. 2003) areupregulated following a period of fat feeding, in line with an
Palatable food mobilizes opioids and dopamine in the re-
increased hunger for fat food. At the same time some satiety
ward system. Opiates and dopamine, when injected into the
MiniReview
Fig. 1a. Hunger and satiety signalling during intake of a standardmeal. Hunger signals, such as ghrelin in the stomach and NPY,
Fig. 1b. Hunger and satiety signalling after a period on a diet of
orexin, AgRP in the hypothalamus, are depressed after intake of
palatable food. Hunger signals are either depressed, like ghrelin in
standard food, while satiety signals like CCK, GLP-1, PYY, insulin
the stomach and NPY in the hypothalamus, in response to a meal
and leptin are raised. Food intake is terminated as a result. For
consisting of palatable food or raised, as for orexin and AgRP in
the hypothalamus. Satiety signals like insulin and leptin are in-creased. Palatable food induces resistance to several satiety signals,documented for CCK, insulin and leptin, resulting in overeating. Food intake is driven by an increased activity in the reward system(dopamine, serotonin and opiates), triggered by the attractivenessof the taste. For abbreviations, see table 1.
nucleus accumbens, will stimulate food intake, in particular
ence is facilitated by factors that enhance the attractiveness
sucrose and fat, thus creating a vicious circle. Such a re-
of palatable food. The attractiveness of food depends not
inforcement mechanism is in line with the view that palat-
only on its taste and content of carbohydrate and fat, but
able food may cause dependence (Gosnell & Krahn 1992;
also on the nutritional state of the individual, i.e. whether
Nestler & Aghajanian 1997). The development of depend-
fasted or well fed (Cabanac & Lafrance 1990; Berridge1991). Long-term food restriction has been shown to aug-ment the rewarding effect not only of food but also of vari-ous drugs of abuse (Cabeza de Vaca & Carr 1998; Carr2002) and intermittent feeding has been shown to increasethe rewarding effect of food (Colantuoni et al. 2002). Notsurprisingly therefore, binge eating can be provoked by foodrestriction (Hagan et al. 2003). A similar sensitizing effecton the reward system by intake restriction is observed foralcohol (Soderpalm & Hansen 1999). Thus food restrictionsensitizes the reward system, which triggers the craving notonly for food (palatable food in particular), but also foraddictive drugs, including alcohol.
With an intermittent feeding protocol, using concen-
trated sugar solutions, sugar dependence has actually been
Fig. 2. Schematic picture of the pathways for appetite signalling
shown to develop in rodents (Colantuoni et al. 2002). Clas-
in the brain during intake of standard food and palatable food,
sically, dependence occurs in two steps. In the first step the
respectively. With standard food, signals are transmitted from the
consumption of the specific item is increased; in the second
gastrointestinal tract to the brain stem, where the energy content
step withdrawal symptoms become manifest in the absence
and taste of the food is registered. In the absence of strongly attract-ive taste, hypothalamus performs the integrated energy sensing
of the consumed item. The withdrawal symptoms include
through the interaction of various appetite signals and their recep-
anxiety, autonomic nervous system abnormalities, and
tors in the hypothalamic nuclei, the arcuate nucleus being one im-
changes in body temperature, tremor and shakes. In an ex-
portant site for regulation of energy balance. Termination of food
periment to study the addictive potential of sugar, rats were
intake occurs. With palatable food the attractive taste of the food
offered a 25% glucose solution together with food pellets 12
is registered, leading to an activation of the reward centre, releasingvarious reward signals, the nucleus accumbens being one important
hr each day (Colantuoni et al. 2002). After one week the
site. The reward centre has close connections to the hypothalamus,
sugar intake had increased three times. Withdrawal of sugar
thereby influencing the hypothalamic energy homeostasis mechan-
precipitated symptoms such as teeth chatter, forepaw
isms. The end result is prolongation of food intake (LH; lateral
tremor and head shakes. Indirect evidence that the opioid
hypothalamus, PVN; paraventricular nucleus, DMH; dorsomedialhypothalamus, VMH; ventromedial hypothalamus). For further ab-
system had been activated during the escalated sugar intake
was provided by the precipitation of withdrawal symptoms
MiniReview
PALATABLE FOOD DISRUPTS APPETITE REGULATION
in response to naloxone (Colantuoni et al. 2002). Anxiety,
References
measured as avoidance behaviour, increased greatly in theserats upon withdrawal of sugar. During the development of
Air, E. L., M. Z. Strowski, S. C. Benoit, S. L. Conarello, G. M.
Salituro, X. M. Guan, K. Liu, S. C. Woods & B. B. Zhang: Small
sugar dependence there was an increased release of dop-
molecule insulin mimetics reduce food intake and body weight
amine and a decreased release of acetylcholine in the nu-
and prevent development of obesity. Nature Med. 2002, 8, 179–
cleus accumbens; withdrawal of sugar reversed the effects
(Colantuoni et al. 2002). Whether man develops depend-
Allen, Y. S., T. E. Adrian, J. M. Allen, K. Tatemoto, T. J. Crow, S.
ence to palatable food and whether withdrawal symptoms
R. Bloom & J. M. Polak: Neuropeptide Y distribution in the rat brain. Science 1983, 221, 877–879.
develop is not known. Further studies are needed to explore
Apfelbaum, M. & A. Mandenoff: Naltrexone suppresses hyper-
the consequences of indulgence in palatable food and to
phagia induced in the rat by a highly palatable diet. Pharmacol.
understand how palatable food affects appetite-regulating
Biochem. Behav. 1981, 15, 89–91.
Appleyard, S. M., M. Hayward, J. I. Young, A. A. Butler, R. D.
Cone, M. Rubinstein & M. J. Low: A role for the endogenous opioid beta-endorphin in energy homeostasis. Endocrinology 2003, 144, 1753–1760. Conclusions
Banks, W. A.: Is obesity a disease of the blood-brain barrier?
In conclusion, appetite regulation involves hunger and satiety
Physiological, pathological, and evolutionary considerations. Curr. Pharm. Des. 2003, 9, 801–809.
signals, released from various peripheral organs to signal to
Banks, W. A., C. R. DiPalma & C. L. Farrell: Impaired transport
the brain. During energy deficiency the expression of hunger
of leptin across the blood-brain barrier in obesity. Peptides 1999,
signals are raised, ghrelin in the stomach and neuropeptide
20, 1341–1345.
Y and the orexins in the hypothalamus. In response to food
Banks, W. A. & C. L. Farrell: Impaired transport of leptin across
the blood-brain barrier in obesity is acquired and reversible.
intake, various satiety signals, e.g. cholecystokinin, gluca-
Amer. J. Physiol. Endocrinol. Metab. 2003, 285, E10–E15.
gon-like peptide 1 and peptide YY (fig. 1a) are released from
Banks, W. A. & A. J. Kastin: Differential permeability of the blood-
the intestine to reach the circulation, signaling to neurones in
brain barrier to two pancreatic peptides: insulin and amylin. Pep-
the brain through vagal afferents. Insulin and leptin are mo-
tides 1998, 19, 883–889.
bilized to induce satiety, adipose tissue releasing leptin in pro-
Bannon, A. W., J. Seda, M. Carmouche, J. M. Francis, M. H. Nor-
man, B. Karbon & M. L. McCaleb: Behavioral characterization
portion to the weight of the fat mass (fig. 1a). After some time
of neuropeptide Y knockout mice. Brain Res. 2000, 868, 79–87.
on a diet consisting of palatable food, the hypothalamic ex-
Baskin, D. G., D. Figlewicz Lattemann, R. J. Seeley, S. C. Woods,
pression of hunger peptides such as neuropeptide Y and the
D. Porte, Jr. & M. W. Schwartz: Insulin and leptin: dual adiposity
orexins is increased. The expression of several satiety signals
signals to the brain for the regulation of food intake and body weight. Brain Res. 1999, 848, 114–123.
is also increased, but the signaling of insulin, leptin and cho-
Batterham, R. L., M. A. Cohen, S. M. Ellis, C. W. Le Roux, D. J.
lecystokinin is blunted or inhibited, promoting over-eating.
Withers, G. S. Frost, M. A. Ghatei & S. R. Bloom: Inhibition of
The continued eating is driven by reward rather than by en-
food intake in obese subjects by peptide YY3–36. N. Engl. J.
ergy deficit (Fig. 1b), resulting in obesity. Med. 2003, 349, 941–948.
Batterham, R. L., M. A. Cowley, C. J. Small, H. Herzog, M. A.
In the brain energy deficit is registered in the hypothala-
Cohen, C. L. Dakin, A. M. Wren, A. E. Brynes, M. J. Low, M.
mus through various nuclei including arcuate nucleus lead-
A. Ghatei, R. D. Cone & S. R. Bloom: Gut hormone PYY(3–
ing to release of hunger signals and activation of their recep-
36) physiologically inhibits food intake. Nature 2002, 418, 650–
tors. Consumption of standard food generates information
Berger, K., U. Sivars, M. S. Winzell, P. Johansson, U. Hellman, C.
on its energy content and taste in the brain stem. This infor-
Rippe & C. Erlanson-Albertsson: Mitochondrial ATP synthase–
mation is transmitted to the hypothalamus to release and/
a possible target protein in the regulation of energy metabolism
or up-regulate various satiety peptides, leading to termin-
in vitro and in vivo. Nutr. Neurosci. 2002, 5, 201–210.
ation of food intake (fig. 2). With palatable food, taste sens-
Berger, K., M. S. Winzell, J. Mei & C. Erlanson-Albertsson: Enter-
ostatin and its target mechanisms during regulation of fat intake.
ing is more intense than with standard food; information is
Physiol. Behav. 2004, 83, 623–630.
transmitted to the reward centre in the nucleus accumbens,
Berridge, K. C.: Modulation of taste affect by hunger, caloric sat-
leading to release and/or up-regulation of reward mediators
iety, and sensory-specific satiety in the rat. Appetite 1991, 16,
like dopamine, serotonin and opiates. The reward centre has
connections with appetite-controlling neurones in the hypo-
Berridge, K. C.: Food reward: brain substrates of wanting and lik-
ing. Neurosci. Biobehav. Rev. 1996, 20, 1–25.
thalamus. With a diet consisting of palatable food, the time
Bi, S., E. E. Ladenheim, G. J. Schwartz & T. H. Moran: A role for
of food intake will be prolonged because of suppressed sat-
NPY overexpression in the dorsomedial hypothalamus in hyper-
iety signaling (fig. 2). This may lead to overeating and adi-
phagia and obesity of OLETF rats. Amer. J. Physiol. Regul.
posity. In the future we will need to find ways to restrain
Integr. Comp. Physiol. 2001, 281, R254–R260.
Billington, C. J., J. E. Briggs, S. Harker, M. Grace & A. S. Levine:
compulsive intake of palatable food. The use of opiate anta-
Neuropeptide Y in hypothalamic paraventricular nucleus: a cent-
gonists as anti-obesity drugs might be considered.
er coordinating energy metabolism. Amer. J. Physiol. 1994, 266, R1765–R1770.
Bittencourt, J. C., F. Presse, C. Arias, C. Peto, J. Vaughan, J. L.
Nahon, W. Vale & P. E. Sawchenko: The melanin-concentrating
The work performed in the author’s laboratory was sup-
hormone system of the rat brain: an immuno- and hybridization
ported by the Swedish Medical Research Council (K2004-
histochemical characterization. J. Comp. Neurol. 1992, 319, 218– MiniReview
Bjorbaek, C., J. K. Elmquist, J. D. Frantz, S. E. Shoelson & J. S.
Davidoff, A. J., M. M. Mason, M. B. Davidson, M. W. Carmody,
Flier: Identification of SOCS-3 as a potential mediator of central
K. K. Hintz, L. E. Wold, D. A. Podolin & J. Ren: Sucrose-in-
leptin resistance. Mol. Cell 1998, 1, 619–625.
duced cardiomyocyte dysfunction is both preventable and revers-
Blum, K., E. R. Braverman, J. M. Holder, J. F. Lubar, V. J. Monas-
ible with clinically relevant treatments. Amer. J. Physiol Endocri-
tra, D. Miller, J. O. Lubar, T. J. Chen & D. E. Comings: Reward
nol. Metab. 2004, 286, E718–E724.
deficiency syndrome: a biogenetic model for the diagnosis and
Del Parigi, A., K. Chen, A. D. Salbe, E. M. Reiman & P. A. Tatar-
treatment of impulsive, addictive, and compulsive behaviors. J.
anni: Are we addicted to food? Obes. Res. 2003, 11, 493–495. Psychoactive Drugs 2000, 32 Suppl, i–iv, 1–112.
Dhillo, W. S., C. J. Small, S. A. Stanley, P. H. Jethwa, L. J. Seal, K.
Blundell, J. E. & A. Gillett: Control of food intake in the obese.
G. Murphy, M. A. Ghatei & S. R. Bloom: Hypothalamic interac-
Obes. Res. 2001, 9, Suppl 4, 263S–270S.
tions between neuropeptide Y, agouti-related protein, cocaine-
Blundell, J. E. & C. L. Lawton: Serotonin and dietary fat intake:
and amphetamine-regulated transcript and alpha-melanocyte-
effects of dexfenfluramine. Metabolism 1995, 44, 33–37.
stimulating hormone in vitro in male rats. J. Neuroendocrinol.
Bray, G. A.: Drug treatment of obesity. Rev. Endocr. Metab. Disord.
2002, 14, 725–730.
2001, 2, 403–418.
Diano, S., B. Horvath, H. F. Urbanski, P. Sotonyi & T. L. Horvath:
Burguera, B., M. E. Couce, G. L. Curran, M. D. Jensen, R. V.
Fasting activates the nonhuman primate hypocretin (orexin) sys-
Lloyd, M. P. Cleary & J. F. Poduslo: Obesity is associated with a
tem and its postsynaptic targets. Endocrinology 2003, 144, 3774–
decreased leptin transport across the blood-brain barrier in rats. Diabetes 2000, 49, 1219–1223.
Dornonville de la Cour, C., M. Bjorkqvist, A. K. Sandvik, I. Bakke,
Buwalda, B., W. A. Blom, J. M. Koolhaas & G. van Dijk: Behav-
C. M. Zhao, D. Chen & R. Hakanson: A-like cells in the rat
ioral and physiological responses to stress are affected by high-
stomach contain ghrelin and do not operate under gastrin con-
fat feeding in male rats. Physiol. Behav. 2001, 73, 371–377.
trol. Regul. Pept. 2001, 99, 141–150.
Cabanac, M. & L. Lafrance: Postingestive alliesthesia: the rat tells
Drewnowski, A., D. D. Krahn, M. A. Demitrack, K. Nairn & B.
the same story. Physiol. Behav. 1990, 47, 539–543.
A. Gosnell: Taste responses and preferences for sweet high-fat
Cabeza de Vaca, S. & K. D. Carr: Food restriction enhances the
foods: evidence for opioid involvement. Physiol. Behav. 1992, 51,
central rewarding effect of abused drugs. J. Neurosci. 1998, 18,
Erickson, J. C., G. Hollopeter & R. D. Palmiter: Attenuation of the
Callahan, H. S., D. E. Cummings, M. S. Pepe, P. A. Breen, C. C.
obesity syndrome of ob/ob mice by the loss of neuropeptide Y
Matthys & D. S. Weigle: Postprandial suppression of plasma
[see comments]. Science 1996, 274, 1704–1707.
ghrelin level is proportional to ingested caloric load but does not
Erlanson-Albertsson, C. & D. York: Enterostatin – a peptide regu-
predict intermeal interval in humans. J. Clin. Endocrinol. Metab.
lating fat intake. Obes. Res. 1997, 5, 360–372.
2004, 89, 1319–1324.
Farooqi, I. S., J. M. Keogh, G. S. Yeo, E. J. Lank, T. Cheetham &
Campfield, L. A., F. J. Smith, Y. Guisez, R. Devos & P. Burn: Re-
S. O’Rahilly: Clinical spectrum of obesity and mutations in the
combinant mouse OB protein: evidence for a peripheral signal
melanocortin 4 receptor gene. New Engl. J. Med. 2003, 348,
linking adiposity and central neural networks. Science 1995, 269,
Farooqi, I. S. & S. O’Rahilly: Monogenic human obesity syn-
Carr, K. D.: Augmentation of drug reward by chronic food restric-
dromes. Recent Prog. Horm. Res. 2004, 59, 409–424.
tion: behavioral evidence and underlying mechanisms. Physiol.
Flatt, P. R., C. J. Bailey, P. Kwasowski & S. K. Swanston-Flatt:
Behav. 2002, 76, 353–364.
Effects of diets rich in sucrose, coconut fat and safflowerseed oil
Challis, B. G., A. P. Coll, G. S. Yeo, S. B. Pinnock, S. L. Dickson,
on the development of the obese hyperglycaemic (ob/ob) syn-
R. R. Thresher, J. Dixon, D. Zahn, J. J. Rochford, A. White, R.
drome in mice. Diabetes Res. 1990, 13, 23–28.
L. Oliver, G. Millington, S. A. Aparicio, W. H. Colledge, A. P.
Frederich, R. C., A. Hamann, S. Anderson, B. Lollmann, B. B.
Russ, M. B. Carlton & S. O’Rahilly: Mice lacking pro-opiomel-
Lowell & J. S. Flier: Leptin levels reflect body lipid content in
anocortin are sensitive to high-fat feeding but respond normally
mice: evidence for diet-induced resistance to leptin action. Nat.
to the acute anorectic effects of peptide-YY (3–36). Proc. Natl.Med. 1995, 1, 1311–1314. Acad. Sci. USA 2004, 101, 4695–4700.
Friedman, J. M. & J. L. Halaas: Leptin and the regulation of body
Cheung, C. C., D. K. Clifton & R. A. Steiner: Proopiomelanocortin
weight in mammals. Nature 1998, 395, 763–770.
neurons are direct targets for leptin in the hypothalamus. Endo-
Froguel, P., B. Guy-Grand & K. Clement: Genetics of obesity: to-
crinology 1997, 138, 4489–4492.
wards the understanding of a complex syndrome. Presse Med.
Clement, K., C. Vaisse, N. Lahlou, S. Cabrol, V. Pelloux, D. Cassu-
2000, 29, 564–571.
to, M. Gourmelen, C. Dina, J. Chambaz, J. M. Lacorte, A. Bas-
Gale, S. M., V. D. Castracane & C. S. Mantzoros: Energy homeo-
devant, P. Bougneres, Y. Lebouc, P. Froguel & B. Guy-Grand: A
stasis, obesity and eating disorders: recent advances in endocrin-
mutation in the human leptin receptor gene causes obesity and
ology. J. Nutr. 2004, 134, 295–298.
pituitary dysfunction. Nature 1998, 392, 398–401.
Gallwitz, B. & W. E. Schmidt: [GLP-1 receptor gen ‘‘knock out’’
Colantuoni, C., P. Rada, J. McCarthy, C. Patten, N. M. Avena, A.
causes glucose intolerance, but no alterations of eating behavior].
Chadeayne & B. G. Hoebel: Evidence that intermittent, excessive
Z. Gastroenterol. 1997, 35, 655–658.
sugar intake causes endogenous opioid dependence. Obes. Res.
Gao, J., L. Ghibaudi, M. van Heek & J. J. Hwa: Characterization
2002, 10, 478–488.
of diet-induced obese rats that develop persistent obesity after 6
Coll, A. P., I. S. Farooqi, B. G. Challis, G. S. Yeo & S. O’Rahilly:
months of high-fat followed by 1 month of low-fat diet. Brain
Proopiomelanocortin and energy balance: insights from human
Res. 2002, 936, 87–90.
and murine genetics. J. Clin. Endocrinol. Metab. 2004, 89, 2557–
Gavrila, A., J. L. Chan, L. C. Miller, K. Heist, N. Yiannakouris &
C. S. Mantzoros: Circulating melanin-concentrating hormone
Corp, E. S., M. Curcio, J. Gibbs & G. P. Smith: The effect of cen-
(MCH), agouti-related protein (AGRP), and alpha melanocyte-
trally administered CCK-receptor antagonists on food intake in
stimulating hormone ({alpha}-MSH) levels in relation to body
rats. Physiol. Behav. 1997, 61, 823–827.
composition; alterations in response to food deprivation and re-
Covasa, M., J. Grahn & R. C. Ritter: High fat maintenance diet
combinant human leptin administration. J. Clin. Endocrinol. Me-
attenuates hindbrain neuronal response to CCK. Regul. Pept.tab. 2005, 90, 1047–1054.
2000, 86, 83–88.
Gerozissis, K., M. Orosco, C. Rouch & S. Nicolaidis: Insulin re-
Cummings, D. E. & M. W. Schwartz: Genetics and pathophysiology
sponses to a fat meal in hypothalamic microdialysates and
of human obesity. Annu. Rev. Med. 2003, 54, 453–471.
plasma. Physiol. Behav. 1997, 62, 767–772. MiniReview
PALATABLE FOOD DISRUPTS APPETITE REGULATION
Giraudo, S. Q., C. M. Kotz, M. K. Grace, A. S. Levine & C. J.
Kim, J. K., R. E. Gimeno, T. Higashimori, H. J. Kim, H. Choi, S.
Billington: Rat hypothalamic NPY mRNA and brown fat un-
Punreddy, R. L. Mozell, G. Tan, A. Stricker-Krongrad, D. J.
coupling protein mRNA after high-carbohydrate or high-fat di-
Hirsch, J. J. Fillmore, Z. X. Liu, J. Dong, G. Cline, A. Stahl, H.
ets. Amer. J. Physiol. 1994, 266, R1578–R1583.
F. Lodish & G. I. Shulman: Inactivation of fatty acid transport
Glass, M. J., E. O’Hare, J. P. Cleary, C. J. Billington & A. S. Levine:
protein 1 prevents fat-induced insulin resistance in skeletal
The effect of naloxone on food-motivated behavior in the obese
muscle. J. Clin. Invest. 2004, 113, 756–763.
Zucker rat. Psychopharmacology (Berl) 1999, 141, 378–384.
Kim, M. S., C. Y. Yoon, K. H. Park, C. S. Shin, K. S. Park, S. Y.
Gosnell, B. A.: Sucrose intake predicts rate of acquisition of cocaine
Kim, B. Y. Cho & H. K. Lee: Changes in ghrelin and ghrelin
self-administration. Psychopharmacology (Berl) 2000, 149, 286–
receptor expression according to feeding status. Neuroreport
2003, 14, 1317–1320.
Gosnell, B. A. & D. D. Krahn: The relationship between saccharin
Klebig, M. L., J. E. Wilkinson, J. G. Geisler & R. P. Woychik:
and alcohol intake in rats. Alcohol. 1992, 9, 203–206.
Ectopic expression of the agouti gene in transgenic mice causes
Grigson, P. S.: Like drugs for chocolate: separate rewards modu-
obesity, features of type II diabetes, and yellow fur. Proc. Natl.
lated by common mechanisms? Physiol. Behav. 2002, 76, 389– Acad. Sci. USA 1995, 92, 4728–4732.
Koob, G. F. & M. Le Moal: Drug abuse: hedonic homeostatic dys-
Gutzwiller, J. P., J. Drewe, S. Ketterer, P. Hildebrand, A. Krau-
regulation. Science 1997, 278, 52–58.
theim & C. Beglinger: Interaction between CCK and a preload
Korner, J., E. Savontaus, S. C. Chua, Jr., R. L. Leibel & S. L.
on reduction of food intake is mediated by CCK-A receptors in
Wardlaw: Leptin regulation of Agrp and Npy mRNA in the rat
humans. Amer. J. Physiol. Regul. Integr. Comp. Physiol. 2000,
hypothalamus. J. Neuroendocrinol. 2001, 13, 959–966. 279, R189–R195.
Krude, H., H. Biebermann & A. Gruters: Mutations in the human
Hagan, M. M., P. C. Chandler, P. K. Wauford, R. J. Rybak & K.
proopiomelanocortin gene. Ann. New Y. Acad. Sci. 2003, 994,
D. Oswald: The role of palatable food and hunger as trigger fac-
tors in an animal model of stress induced binge eating. Int. J.
Krugel, U., T. Schraft, H. Kittner, W. Kiess & P. Illes: Basal and
Eat. Disord. 2003, 34, 183–197.
feeding-evoked dopamine release in the rat nucleus accumbens is
Hagan, M. M., F. D. Holguin, C. E. Cabello, D. R. Hanscom &
depressed by leptin. Eur. J. Pharmacol. 2003, 482, 185–187.
D. E. Moss: Combined naloxone and fluoxetine on deprivation-
Lam, W. F., H. A. Gielkens, S. Y. de Boer, C. B. Lamers & A. A.
induced binge eating of palatable foods in rats. Pharmacol. Bio-
Masclee: Influence of hyperglycemia on the satiating effect of
chem. Behav. 1997, 58, 1103–1107.
CCK in humans. Physiol. Behav. 1998, 65, 505–511.
Halford, J. C. & J. E. Blundell: Separate systems for serotonin and
Lawton, C. L., J. K. Wales, A. J. Hill & J. E. Blundell: Seroton-
leptin in appetite control. Ann. Med. 2000, 32, 222–232.
inergic manipulation, meal-induced satiety and eating pattern:
Helm, K. A., P. Rada & B. G. Hoebel: Cholecystokinin combined
effect of fluoxetine in obese female subjects. Obes. Res. 1995, 3,
with serotonin in the hypothalamus limits accumbens dopamine
release while increasing acetylcholine: a possible satiation mech-
Le Magnen, J., P. Marfaing-Jallat, D. Miceli & M. Devos: Pain
anism. Brain Res. 2003, 963, 290–297.
modulating and reward systems: a single brain mechanism? Phar-
Herve, C. & D. Fellmann: Changes in rat melanin-concentrating
macol. Biochem. Behav. 1980, 12, 729–733.
hormone and dynorphin messenger ribonucleic acids induced by
Leibel, R. L.: The role of leptin in the control of body weight. Nutr.
food deprivation. Neuropeptides 1997, 31, 237–242. Rev. 2002, 60, S15–S19; discussion S68–84, 85–87.
Huang, X. F., M. Han, T. South & L. Storlien: Altered levels of
Levine, A. S., C. M. Kotz & B. A. Gosnell: Sugars and fats: the
POMC, AgRP and MC4-R mRNA expression in the hypothala-
neurobiology of preference. J. Nutr. 2003, 133, 831S–834S.
mus and other parts of the limbic system of mice prone or resis-
Lin, S., L. H. Storlien & X. F. Huang: Leptin receptor, NPY,
tant to chronic high-energy diet-induced obesity. Brain Res. 2003,
POMC mRNA expression in the diet-induced obese mouse brain. 992, 9–19. Brain Res. 2000, 875, 89–95.
Huang, X. F., X. Xin, P. McLennan & L. Storlien: Role of fat
Loos, R. J. & C. Bouchard: Obesity–is it a genetic disorder? J. In-
amount and type in ameliorating diet-induced obesity: insights
tern. Med. 2003, 254, 401–425.
at the level of hypothalamic arcuate nucleus leptin receptor,
Lord, J. A., A. A. Waterfield, J. Hughes & H. W. Kosterlitz: Endog-
neuropeptide Y and pro-opiomelanocortin mRNA expression.
enous opioid peptides: multiple agonists and receptors. NatureDiabetes Obes. Metab. 2004, 6, 35–44.
1977, 267, 495–499.
Ito, M., A. Gomori, A. Ishihara, Z. Oda, S. Mashiko, H. Matsushi-
Maffei, M., J. Halaas, E. Ravussin, R. E. Pratley, G. H. Lee, Y.
ta, M. Yumoto, H. Sano, S. Tokita, M. Moriya, H. Iwaasa & A.
Zhang, H. Fei, S. Kim, R. Lallone, S. Ranganathan, et al.: Leptin
Kanatani: Characterization of MCH-mediated obesity in mice.
levels in human and rodent: measurement of plasma leptin and
Amer. J. Physiol. Endocrinol. Metab. 2003, 284, E940–E945.
ob RNA in obese and weight-reduced subjects. Nature Med.
Kaga, T., A. Inui, M. Okita, A. Asakawa, N. Ueno, M. Kasuga,
1995, 1, 1155–1161.
M. Fujimiya, N. Nishimura, R. Dobashi, Y. Morimoto, I. M.
Moesgaard, S. G., B. Ahren, R. D. Carr, D. X. Gram, C. L. Brand &
Liu & J. T. Cheng: Modest overexpression of neuropeptide Y in
F. Sundler: Effects of high-fat feeding and fasting on ghrelin ex-
the brain leads to obesity after high-sucrose feeding. Diabetes
pression in the mouse stomach. Regul. Pept. 2004, 120, 261–267.
2001, 50, 1206–1210.
Montague, C. T., I. S. Farooqi, J. P. Whitehead, M. A. Soos, H.
Kawai, K., K. Sugimoto, K. Nakashima, H. Miura & Y. Ninomiya:
Rau, N. J. Wareham, C. P. Sewter, J. E. Digby, S. N. Mohammed,
Leptin as a modulator of sweet taste sensitivities in mice. Proc.
J. A. Hurst, C. H. Cheetham, A. R. Earley, A. H. Barnett, J. B. Natl. Acad. Sci. USA 2000, 97, 11044–11049.
Prins & S. O’Rahilly: Congenital leptin deficiency is associated
Kelley, A. E.: Ventral striatal control of appetitive motivation: role
with severe early-onset obesity in humans. Nature 1997, 387, 903–
in ingestive behavior and reward-related learning. Neurosci. Bi-obehav. Rev. 2004, 27, 765–776.
Moraes, R. C., A. Blondet, K. Birkenkamp-Demtroeder, J. Tirard,
Kelley, A. E., V. P. Bakshi, S. N. Haber, T. L. Steininger, M. J.
T. F. Orntoft, A. Gertler, P. Durand, D. Naville & M. Begeot:
Will & M. Zhang: Opioid modulation of taste hedonics within
Study of the alteration of gene expression in adipose tissue of
the ventral striatum. Physiol. Behav. 2002, 76, 365–377.
diet-induced obese mice by microarray and reverse transcription-
Kim, E. M., C. C. Welch, M. K. Grace, C. J. Billington & A. S.
polymerase chain reaction analyses. Endocrinology 2003, 144,
Levine: Chronic food restriction and acute food deprivation de-
crease mRNA levels of opioid peptides in arcuate nucleus. Amer.
Moran, T. H., A. R. Baldessarini, C. F. Salorio, T. Lowery & G. J. J. Physiol. 1996, 270, R1019–R1024.
Schwartz: Vagal afferent and efferent contributions to the inhi-
MiniReview
bition of food intake by cholecystokinin. Amer. J. Physiol. 1997,
Baskin: Central nervous system control of food intake. Nature272, R1245–R1251.
2000, 404, 661–671.
Naslund, E., B. Barkeling, N. King, M. Gutniak, J. E. Blundell, J.
Segal-Lieberman, G., R. L. Bradley, E. Kokkotou, M. Carlson, D.
J. Holst, S. Rossner & P. M. Hellstrom: Energy intake and appe-
J. Trombly, X. Wang, S. Bates, M. G. Myers, Jr., J. S. Flier &
tite are suppressed by glucagon-like peptide-1 (GLP-1) in obese
E. Maratos-Flier: Melanin-concentrating hormone is a critical
men. Int. J. Obes. Relat. Metab. Disord. 1999, 23, 304–311.
mediator of the leptin-deficient phenotype. Proc. Natl. Acad. Sci.
Nesse, R. M. & K. C. Berridge: Psychoactive drug use in evolution-
USA 2003, 100, 10085–10090.
ary perspective. Science 1997, 278, 63–66.
Shearman, L. P., R. E. Camacho, D. Sloan Stribling, D. Zhou, M.
Nestler, E. J. & G. K. Aghajanian: Molecular and cellular basis of
A. Bednarek, D. L. Hreniuk, S. D. Feighner, C. P. Tan, A. D.
addiction. Science 1997, 278, 58–63.
Howard, L. H. Van der Ploeg, D. E. MacIntyre, G. J. Hickey &
Niimi, M., M. Sato & T. Taminato: Neuropeptide Y in central con-
A. M. Strack: Chronic MCH-1 receptor modulation alters appe-
trol of feeding and interactions with orexin and leptin. Endocrine
tite, body weight and adiposity in rats. Eur. J. Pharmacol. 2003,
2001, 14, 269–273. 475, 37–47.
Ookuma, K., C. Barton, D. A. York & G. A. Bray: Effect of enter-
Shi, Y.: Beyond skin color: emerging roles of melanin-concentrating
ostatin and kappa-opioids on macronutrient selection and con-
hormone in energy homeostasis and other physiological func-
sumption. Peptides 1997, 18, 785–791.
tions. Peptides 2004, 25, 1605–1611.
Owji, A. A., Z. Khoshdel, F. Sanea, M. R. Panjehshahin, M. Sho-
Shigemura, N., R. Ohta, Y. Kusakabe, H. Miura, A. Hino, K. Koy-
jaee Fard, D. M. Smith, H. A. Coppock, M. A. Ghatei & S. R.
ano, K. Nakashima & Y. Ninomiya: Leptin modulates behavioral
Bloom: Effects of intracerebroventricular injection of glucagon
responses to sweet substances by influencing peripheral taste
like peptide-1 and its related peptides on serotonin metabolism
structures. Endocrinology 2004, 145, 839–847.
and on levels of amino acids in the rat hypothalamus. Brain Ress
Sipols, A. J., J. Bayer, R. Bennett & D. P. Figlewicz: Intraventricular
2002, 929, 70–75.
insulin decreases kappa opioid-mediated sucrose intake in rats.
Park, M., L. Lin, S. Thomas, H. D. Braymer, P. M. Smith, D. H. Peptides 2002, 23, 2181–2187.
Harrison & D. A. York: The F1-ATPase beta-subunit is the puta-
Smith, G. P. & J. Gibbs: Cholecystokinin: a putative satiety signal.
tive enterostatin receptor. Peptides 2004, 25, 2127–2133. Pharmacol. Biochem. Behav. 1975, 3, 135–138.
Pelchat, M.: Of human bondage: Food craving, obsession, compul-
So¨derpalm, A. H. & S. Hansen: Alcohol alliesthesia: food restric-
sion and addiction. Physiol. & Behav. 2002, 76, 347–352.
tion increases the palatability of alcohol through a corticos-
Pert, C. B. & S. H. Snyder: Opiate receptor: demonstration in ner-
terone-dependent mechanism. Physiol. Behav. 1999, 67, 409–
vous tissue. Science 1973, 179, 1011–1014.
Polson, D. A. & M. P. Thompson: Adiponutrin mRNA expression
Stanley, B. G. & S. F. Leibowitz: Neuropeptide Y injected in the
in white adipose tissue is rapidly induced by meal-feeding a high-
paraventricular hypothalamus: a powerful stimulant of feeding
sucrose diet. Biochem. Biophys. Res. Commun. 2003, 301, 261–
behavior. Proc. Natl. Acad. Sci. USA 1985, 82, 3940–3943.
Steinberg, G. R., A. C. Smith, S. Wormald, P. Malenfant, C. Col-
Qu, D., D. S. Ludwig, S. Gammeltoft, M. Piper, M. A. Pelleymount-
lier & D. J. Dyck: Endurance training partially reverses dietary-
er, M. J. Cullen, W. F. Mathes, R. Przypek, R. Kanarek & E.
induced leptin resistance in rodent skeletal muscle. Amer. J. Phy-
Maratos-Flier: A role for melanin-concentrating hormone in the
siol. Endocrinol. Metab. 2004, 286, E57–E63.
central regulation of feeding behaviour. Nature 1996, 380, 243–
Strubbe, J. H., D. Porte, Jr. & S. C. Woods: Insulin responses and
glucose levels in plasma and cerebrospinal fluid during fasting
Rada, P., G. P. Mark & B. G. Hoebel: Galanin in the hypothalamus
and refeeding in the rat. Physiol. Behav. 1988, 44, 205–208.
raises dopamine and lowers acetylcholine release in the nucleus
Sun, G., Z. Tian, T. Murata, K. Narita, K. Honda & T. Higuchi:
accumbens: a possible mechanism for hypothalamic initiation of
Central and peripheral immunoreactivity of melanin-concentrat-
feeding behavior. Brain Res. 1998, 798, 1–6.
ing hormone in hypothalamic obese and lactating rats. J. Neuro-
Raposinho, P. D., D. D. Pierroz, P. Broqua, R. B. White, T. Pedrazzi-
endocrinol. 2004, 16, 79–83.
ni & M. L. Aubert: Chronic administration of neuropeptide Y
Szczypka, M. S., M. A. Rainey & R. D. Palmiter: Dopamine is
into the lateral ventricle of C57BL/6J male mice produces an
required for hyperphagia in Lep(ob/ob) mice. Nat. Genet. 2000,
obesity syndrome including hyperphagia, hyperleptinemia, insu-
25, 102–104.
lin resistance, and hypogonadism. Mol. Cell Endocrinol. 2001,
Tartaglia, L. A., M. Dembski, X. Weng, N. Deng, J. Culpepper, R. 185, 195–204.
Devos, G. J. Richards, L. A. Campfield, F. T. Clark, J. Deeds &
Rossi, M., S. J. Choi, D. O’Shea, T. Miyoshi, M. A. Ghatei & S. R. et al.: Identification and expression cloning of a leptin receptor,
Bloom: Melanin-concentrating hormone acutely stimulates feed-
OB-R. Cell 1995, 83, 1263–1271.
ing, but chronic administration has no effect on body weight.
Taubes, G.: As obesity rates rise, experts struggle to explain why. Endocrinology 1997, 138, 351–355. Science 1998, 280, 1367–1368.
Sakurai, T., A. Amemiya, M. Ishii, I. Matsuzaki, R. M. Chemelli,
Tritos, N. A. & E. Maratos-Flier: Two important systems Clark, J.
H. Tanaka, S. C. Williams, J. A. Richarson, G. P. Kozlowski, S.
Deeds, et al.: Identification and expression cloning trating hor-
Wilson, J. R. Arch, R. E. Buckingham, A. C. Haynes, S. A. Carr,
mone. Neuropeptides 1999, 33, 339–349.
R. S. Annan, D. E. McNulty, W. S. Liu, J. A. Terrett, N. A.
Tschop, M., D. B. Flora, J. P. Mayer & M. L. Heiman: Hypophysec-
Elshourbagy, D. J. Bergsma & M. Yanagisawa: Orexins and orex-
tomy prevents ghrelin-induced adiposity and increases gastric
in receptors: a family of hypothalamic neuropeptides and G pro-
ghrelin secretion in rats. Obes. Res. 2002, 10, 991–999.
tein-coupled receptors that regulate feeding behavior. Cell 1998,
Tschop, M., D. L. Smiley & M. L. Heiman: Ghrelin induces adi-
92, 573–585.
posity in rodents. Nature 2000, 407, 908–913.
Saper, C. B., T. C. Chou & J. K. Elmquist: The need to feed: homeo-
Tschop, M., C. Weyer, P. A. Tataranni, V. Devanarayan, E. Ravus-
static and hedonic control of eating. Neuron. 2002, 36, 199–
sin & M. L. Heiman: Circulating ghrelin levels are decreased in
human obesity. Diabetes 2001, 50, 707–709.
Schwartz, M. W., J. L. Marks, A. J. Sipols, D. G. Baskin, S. C.
Turton, M. D., D. O’Shea, I. Gunn, S. A. Beak, C. M. Edwards, K.
Woods, S. E. Kahn & D. Porte, Jr.: Central insulin administration
Meeran, S. J. Choi, G. M. Taylor, M. M. Heath, P. D. Lambert,
reduces neuropeptide Y mRNA expression in the arcuate nucleus
J. P. Wilding, D. M. Smith, M. A. Ghatei, J. Herbert & S. R.
of food-deprived lean (Fa/Fa) but not obese (fa/fa) Zucker rats.
Bloom: A role for glucagon-like peptide-1 in the central regula-
Endocrinology 1991, 128, 2645–2647.
tion of feeding. Nature 1996, 379, 69–72.
Schwartz, M. W., S. C. Woods, D. Porte, Jr., R. J. Seeley & D. G.
Vaccarino, F. J.: Nucleus accumbens dopamine-CCK interactions in
MiniReview
PALATABLE FOOD DISRUPTS APPETITE REGULATION
psychostimulant reward and related behaviors. Neurosci. Bi-
anisms of the hypophysiotropic action of ghrelin. Neuroendocrin-obehav. Rev. 1994, 18, 207–214. ology 2002, 76, 316–324.
van Citters, G. W., M. Kabir, S. P. Kim, S. D. Mittelman, M. K.
Wurtman, R. J. & J. J. Wurtman: Brain serotonin, carbohydrate-
Dea, P. L. Brubaker & R. N. Bergman: Elevated glucagon-like
craving, obesity and depression. Obes. Res. 1995, 3, Suppl 4,
peptide-1-(7–36)-amide, but not glucose, associated with hyper-
insulinemic compensation for fat feeding. J. Clin. Endocrinol.
Yeo, G. S., I. S. Farooqi, S. Aminian, D. J. Halsall, R. G. Stan-
Metab. 2002, 87, 5191–5198.
hope & S. O’Rahilly: A frameshift mutation in MC4R associated
Van Ree, J. M., R. J. Niesink, L. Van Wolfswinkel, N. F. Ramsey,
with dominantly inherited human obesity. Nat. Genet. 1998, 20,
M. M. Kornet, W. R. Van Furth, L. J. Vanderschuren, M. A.
Gerrits & C. L. Van den Berg: Endogenous opioids and reward.
Yeo, G. S., E. J. Lank, I. S. Farooqi, J. Keogh, B. G. Challis & S. Eur. J. Pharmacol. 2000, 405, 89–101.
O’Rahilly: Mutations in the human melanocortin-4 receptor gene
Wang, G. J., N. D. Volkow, J. Logan, N. R. Pappas, C. T. Wong, W.
associated with severe familial obesity disrupts receptor function
Zhu, N. Netusil & J. S. Fowler: Brain dopamine and obesity.
through multiple molecular mechanisms. Hum. Mol. Genet. 2003,
Lancet 2001, 357, 354–357. 12, 561–574.
Welch, C. C., E. M. Kim, M. K. Grace, C. J. Billington & A. S.
Yeomans, M. R. & R. W. Gray: Selective effects of naltrexone on
Levine: Palatability-induced hyperphagia increases hypothalamic
food pleasantness and intake. Physiol. Behav. 1996, 60, 439–446.
Dynorphin peptide and mRNA levels. Brain Res. 1996, 721, 126–
Yeomans, M. R., R. W. Gray, C. J. Mitchell & S. True: Independent
effects of palatability and within-meal pauses on intake and
Williams, L. M., C. L. Adam, J. G. Mercer, K. M. Moar, D. Slater,
appetite ratings in human volunteers. Appetite 1997, 29, 61–76.
L. Hunter, P. A. Findlay & N. Hoggard: Leptin receptor and
Zamir, N., G. Skofitsch & D. M. Jacobowitz: Distribution of im-
neuropeptide Y gene expression in the sheep brain. J. Neuroen-
munoreactive melanin-concentrating hormone in the central ner-
docrinol. 1999, 11, 165–169.
vous system of the rat. Brain Res. 1986, 373, 240–245.
Woods, S. C., M. W. Schwartz, D. G. Baskin & R. J. Seeley: Food
Zhang, M., C. Balmadrid & A. E. Kelley: Nucleus accumbens
intake and the regulation of body weight. Annu. Rev. Psychol.
opioid, GABaergic, and dopaminergic modulation of palatable
2000, 51, 255–277.
food motivation: contrasting effects revealed by a progressive
Woods, S. C., R. J. Seeley, D. G. Baskin & M. W. Schwartz: Insulin
ratio study in the rat. Behav. Neurosci. 2003, 117, 202–211.
and the blood-brain barrier. Curr. Pharm. Des. 2003, 9, 795–800.
Zhang, M. & A. E. Kelley: Opiate agonists microinjected into the
Wortley, K. E., G. Q. Chang, Z. Davydova & S. F. Leibowitz: Pep-
nucleus accumbens enhance sucrose drinking in rats. Psychophar-
tides that regulate food intake: orexin gene expression is increased
macology (Berl.) 1997, 132, 350–360.
during states of hypertriglyceridemia. Amer. J. Physiol. Regul.
Zhang, M. & A. E. Kelley: Intake of saccharin, salt, and ethanol
Integr. Comp. Physiol. 2003, 284, R1454–R1465.
solutions is increased by infusion of a mu opioid agonist into the
Wren, A. M., L. J. Seal, M. A. Cohen, A. E. Brynes, G. S. Frost,
nucleus accumbens. Psychopharmacology (Berl.) 2002, 159, 415–
K. G. Murphy, W. S. Dhillo, M. A. Ghatei & S. R. Bloom: Ghrel-
in enhances appetite and increases food intake in humans. J. Clin.
Zigman, J. M. & J. K. Elmquist: Minireview: From anorexia to
Endocrinol. Metab. 2001, 86, 5992.
obesity – the yin and yang of body weight control. Endocrinology
Wren, A. M., C. J. Small, C. V. Fribbens, N. M. Neary, H. L. Ward,
2003, 144, 3749–3756.
L. J. Seal, M. A. Ghatei & S. R. Bloom: The hypothalamic mech-
Relaciones entre estructura e historia de la ciencia Dr. César Lorenzano Este artículo fue presentado en el Primer Congreso Latinoamericano de Historia de la Ciencia y la Tecnología que se realizó en Puebla, México, en 1982, y en el que se fundó la Sociedad Latinoamericana de Historia de la Ciencia y la Tecnología. Se encuentra publicado en la primera edición (1988) de La estruct
FOR IMMEDIATE RELEASE HUMAN RIGHTS AUTHORITY - NORTHWEST REGION Case Summary: the HRA found no additional violations. The Human Rights Authority (HRA) opened an investigation after receiving complaints of possible rights violations at Hammett House, a sixteen-bed intermediate care facility for persons with developmental disabilities in Sterling. The home is managed by Frances House, a n
 C Basic & Clinical Pharmacology & Toxicology 2005, 97, 61–73.
C Basic & Clinical Pharmacology & Toxicology 2005, 97, 61–73.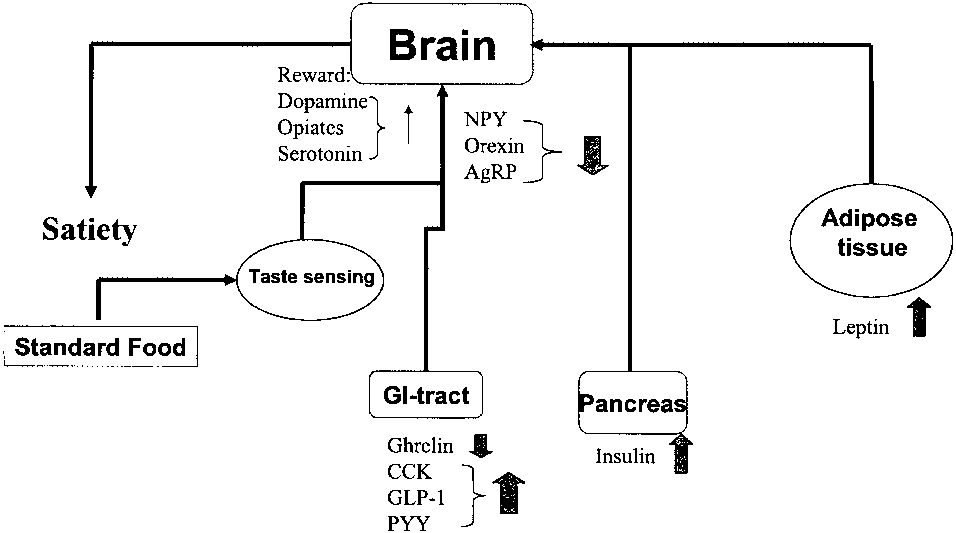
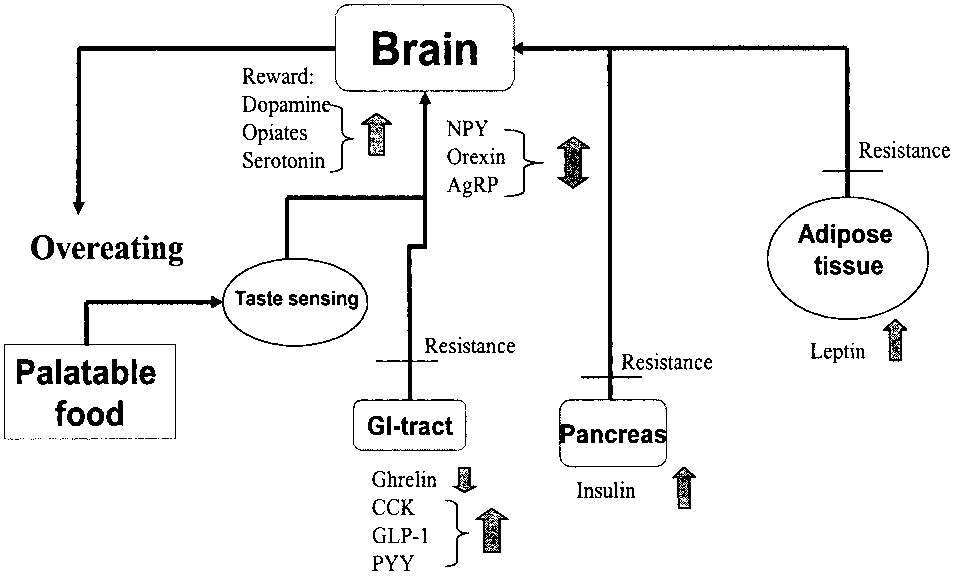
 MiniReview
MiniReview