Levitra has a minimal amount of contraindications which has increased its popularity levitra uk You can buy quality certified medications from us at an affordable price.
Ubik.microbiol.washington.edu
Human Immunodeficiency Virus Type 1 Subtypes Differ in Disease Progression Phyllis J. Kanki, Donald J. Hamel, Jean-Louis Sankale´, Departments of Immunology and Infectious Diseases and HarvardChung-cheng Hsieh, Ibou Thior, Francis Barin, AIDS Institute, Harvard School of Public Health, Boston, andStephen A. Woodcock, Aı¨ssatou Gue`ye-Ndiaye, University of Massachusetts Cancer Center, Worcester, Massachusetts;Laboratory of Virology, University of Tours, Tours, France;Er Zhang, Monty Montano, Tidiane Siby, Laboratory of Bacteriology and Virology, Cheikh Anta DiopRichard Marlink, Ibrahima NDoye, Myron E. Essex, University, and Institut Hygiene Sociale, Dakar, Senegaland Souleymane MBoup At least 10 different genetic human immunodeficiency virus type 1 (HIV-1) subtypes (A–J) are responsible for the AIDS pandemic. Much of the understanding of HIV-1 disease pro- gression derives from studies in the developed world where HIV infection is almost exclusively subtype B. This has led many to question whether the properties and consequences of HIV- 1 infection can be generalized across subtypes that afflict the majority of infected persons in the developing world. From 1985 to 1997, a prospective study of registered female sex workers in Senegal tracked the introduction and spread of HIV-1 subtypes A, C, D, and G. In clinical follow-up, the AIDS-free survival curves differed by HIV-1 subtype. Women infected with a non-A subtype were 8 times more likely to develop AIDS than were those infected with subtype A (hazard ratio ϭ 8.23; P ϭ .009), the predominant subtype in the study. These data suggest that HIV-1 subtypes may differ in rates of progression to AIDS.
The human immunodeficiency virus (HIV) type 1 epidemic
transmission potentials. This prospective study was undertaken
in many parts of sub-Saharan Africa is at least several decades
to determine the dynamics of various HIV-1 subtype infections
old, considerably older than in other parts of the world, such
in a high-risk population over a decade of observation.
as Asia. HIV burden for the African continent is estimated at
The recognized differences in transmission and virulence of
15–20 million, with heterosexual and perinatal transmission the
HIV-2, compared with HIV-1, indicate that HIV viruses can
major routes of infection [1]. Most of the 10 known HIV-1
have different pathogenic potentials. Prospective studies that
subtypes have been reported in sub-Saharan Africa, although
identified and followed seroincident infection were critical to
surveillance data on subtypes G, H, I, and J remain preliminary
the observation that HIV-2 was less transmissible and slower
[2–5]. Current information on the prevalence of HIV-1 subtypes
in disease progression than HIV-1 [6–8]. This lends support to
is cross-sectional, frequently based on small numbers of samples
the hypothesis that genetically distinct subtypes of HIV-1 might
that were readily available, and often derived from patients at
also differ in their biologic properties. Subtype differences in
different times during their HIV infection [2]. Thus, although
transmission and disease potential would drastically alter our
studies in certain African countries have described multiple
predictions of the future of the HIV epidemic and its impact.
HIV-1 subtypes, it is not known whether subtypes entered these
Estimates of the time to development of AIDS after HIV
populations at different times or whether the distribution of
infection are long, variable, and based primarily on data from
subtypes reflects the dynamics of different subtype-specific
US and European studies with subtype B–infected persons. Although few data are available from the developing world,where non-B subtypes predominate, cross-sectional studies sug-
Received 4 May 1998; revised 25 August 1998.
gest that HIV non-B subtypes are common in AIDS patients
Presented in part: 12th World AIDS Conference, Geneva, Switzerland,
[9, 10]. However, differences in pathogenicity between HIV sub-
28 June–3 July 1998 (abstract 277).
Informed consent was obtained from all study participants. The protocol
types might be best discerned from prospective studies, where
was approved by ethical review committees in Dakar, Senegal, and at Har-
the time to AIDS development can be estimated and compared.
vard School of Public Health. Human experimentation guidelines of the US
Given the known differences in the geographic distribution and
Department of Health and Human Services were followed.
Financial support: US Army Medical Research and Materiel Command
associated epidemics of different HIV subtypes, we asked
(grants 17-87-7072, 17-90-C-0138, 17-95-C-5005) and NIH (grants AI-
whether HIV-1 subtypes might differ in their rates of disease
30795, AI-35173). The study was conducted under the Inter-University
development. This study is the first to evaluate disease pro-
Convention for the Prevention of AIDS and Other Sexually TransmittedDiseases. I.T., A.G.-N. and J.-L.S. are Fogarty International fellows.
gression of different HIV-1 subtypes in a cohort of prospectively
Reprints or correspondence: Dr. P. J. Kanki, Dept. of Immunology and
followed persons with known time of infection.
Infectious Diseases, Harvard School of Public Health, 651 Huntington Ave.,Boston, MA 02115 ([email protected]). The Journal of Infectious Diseases 1999; 179:68–73
Since 1985, all women registered as prostitutes at the Institut
1999 by the Infectious Diseases Society of America. All rights reserved.
d’Hygie`ne Sociale clinic in Dakar, Senegal, have been serologically
screened for exposure to HIV-1 and HIV-2. The clinic provides
Multiple alignments were done with the Clustal package (W 1.6)
clinical examinations and treatments of sexually transmitted dis-
with minor manual adjustment when necessary [14]. Phylogenetic
eases (STDs) during biannual visits, which are required for the
analysis was performed by the neighbor-joining method, and re-
legal registration of prostitutes. All women who gave informed
liability was estimated from 1000 bootstrap resamplings. Repre-
consent were enrolled in the study, and a baseline questionnaire
sentative sequences of various HIV subtypes A–I were included in
and health evaluation was administered by study physicians. The
the analysis [2]. The nucleotide sequences were submitted to
dynamic nature of this clinic resulted in an open cohort design in
GenBank (accession numbers: AF085284–085327 and AF020819–
which subjects were continuously identified and enrolled over the
study period. The study design, recruitment procedures, and var-
In samples where envelope sequences could not be amplified,
ious protocols have been described [6–8, 11].
nested PCR in the long-terminal-repeat (LTR) region was done for
At the time of enrollment and at subsequent biannual visits, a
subtype assignment. Oligonucleotide primers were designed on the
complete physical examination was done that emphasized HIV-
basis of an analysis of conserved regions within the LTR across
related signs and symptoms. All biannual blood samples were tested
the clades represented in the Los Alamos (NM) National Labo-
for antibodies to each virus by immunoblot for HIV-1 and HIV-2
ratory HIV database (A–G, O). Upstream primers for the LTR
antibodies in the Dakar and Boston laboratories. T cell subsets
were PH5KPN. Downstream primers for the LTR were PH3 LTR
were determined annually, and DNA was extracted from the pe-
ripheral blood mononuclear cells that remained after T cell subset
PH3 LTR: 5 -CTGAGGGATCTCTAGTTACCAGAG-3 ; PH56:
analysis by proteinase K–phenol-chloroform extraction.
5 -ATTGAGGCTTAAGCAGTGGG-3 ). Samples were subjected
Seroincident HIV-1 cases were defined as women who were pre-
to 30–39 amplification cycles consisting of a denaturing step (94ЊC,
viously HIV-negative and subsequently seroconverted to HIV-1.
30 s), a primer annealing step (50ЊC, 30 s), and a primer extension
The time of infection, or seroconversion date, was estimated as the
midpoint between the last seronegative and first seropositive sam-ples. HIV-1–positive person-years of observation was calculatedfrom the seroconversion date until the AIDS disease outcome.
AIDS was diagnosed by the CDC revised surveillance case defi-
From February 1985 to December 1995, 1683 seronegative
nition, which includes !200 CD4 cells [12]. Cases were censored at
women were enrolled in the study and followed; 81 women
their last clinic examination date or date of death (not caused byAIDS); the observation period ended on 1 January 1997. Kaplan-
subsequently seroconverted to HIV-1. DNA samples were avail-
Meier AIDS-free survival analysis and Cox proportional hazards
able from 54 of the 81 HIV-1 seroconverters, and HIV-1 subtype
models for AIDS outcome were done as described (Stata, College
was determined. HIV-1 subtype was assigned based on the C2-
V3 sequence in 53 of 54 samples; LTR sequences were used for
HIV-1 subtype was determined by polymerase chain reaction
the remaining sample to determine subtype after the envelope
(PCR) amplification of ∼350 bp in the HIV-1 C2–V3 envelope
sequences failed to provide subtype information. Subtypes were
region. A nested PCR was done using two sets of primers: WT1,
determined by phylogenetic tree analysis; based on 1000 boot-
5 -GCTGGTTTTGCGATTCTAAAGTGTA (6884–6908, posi-
strap resamplings, high bootstrap values demonstrated the
tions relative to HXB2) and WT2, 5 -CAATAGAAAAATTCC-
branching order of sequences with known HIV-1 subtypes A
CCTCCACAAT (7353–7377), for the first round; and published
(93.4%), C (96.4%), D (92.1%), and G (99.0%) (figure 1).
primers, KK30–KK40, for the second round [13].
Subtype A was found in 37 (68.5%) of 54 seroconverters,
The PCR reactions were in a 100-mL volume reaction mixture
subtype C in 5 (9.3%), subtype D in 5 (9.3%), and subtype G
containing 1 mg of DNA, 10 mL of 10ϫ PCR buffer II and 2.5 Uof Taq polymerase (Perkin-Elmer PCR Reagents, Roche Molecular
in 6 (11.1%). One (1.8%) of the V3 envelope sequences had
Systems, Branchburg, NJ), 0.2 mM of each dideoxy-nucleotide, 2
significant variation from other subtype A sequences. Exami-
mM MgCl , and 20 pmol of each primer. Each reaction was sub-
nation by the Recombinant Identification Program [15] re-
jected to 30–35 cycles of denaturation (94ЊC, 45 s), annealing (45
vealed a mosaic virus composed of subtypes A and G, which
s at 57ЊC and 55ЊC in the first and second rounds, respectively),
was further verified by sequencing of the complete gp120 por-
and extension (72ЊC, 45 s), followed by a final extension (72ЊC, 3
tion of the envelope. Most of the gp120 aligned with subtype
min) in an automatic thermal cycler. Negative controls included
A, but ∼300 bp in the V2–C2 region aligned with subtype G.
deionized water and negative peripheral blood lymphocyte DNA
This woman was infected in 1994 and is currently healthy; she
was not included in further analyses because of the mosaic
The PCR product was purified and directly sequenced using the
virus. While HIV-1 subtype assignment utilizing the V3 enve-
second-round primers (KK30 and KK40). When necessary, the
lope sequence has high specificity and sensitivity, it is inade-
purified product was cloned in pCR2.1 vector (T/A cloning; In-
quate to address the potential existence of recombinant or mo-
vitrogen, San Diego). Sequences were determined by dye termi-nator cycle sequencing using Taq polymerase and an automatic
saic viruses [16, 17]. Thus, it is possible that other women in
sequencer ABI 373A (both Perkin-Elmer, Applied Biosystem Di-
our study harbor viruses that are monotypic within the V3
vision, Foster City, CA). In all cases, sequences were determined
region but mosaic or recombinant in other regions of the
for both strands of DNA, and in many instances multiple clones
The distribution of seroconverters over the 10-year period is
Figure 1.
Phylogenetic tree analysis by neighbor-joining method of V3 envelope sequences from registered female sex workers who seroconverted
between 1985 and mid-1995. Representative V3 envelope sequences of various HIV-1 subtypes A–I are indicated in underlined bold text [2]. Alignment was subjected to 1000 bootstrap resamplings; no. of resamplings is indicated at major branch points. Horizontal distance correspondsto genetic relatedness.
illustrated by HIV subtype designated at the estimated sero-conversion date (figure 2). In this high-risk group of women,the first HIV-1 seroconversions occurred in late 1987. Through-out the study period, subtype A was the most common HIV-1 subtype. The first seroconversion to subtype C occurred latein 1987; the last C seroconversion was observed in late 1993. Similarly, subtype D was observed early in 1987 and then spo-radically until early 1994. Subtype G infections first appearedin mid-1992 and in subsequent years represented 20% of theincident HIV-1 subtypes.
We are cautious in our interpretation of the subtype dynamics
in this cohort, since DNA samples were unavailable for 27 ofthe 81 HIV-1 seroconverters observed in the study. A logisticregression analysis, including age at registration, years of reg-istered prostitution, nationality, and year of seroconversion,showed that women of Ghanaian nationality were less likelyto be subtyped (P ϭ .02). Ghanaian women represented a smallfraction of all HIV-1 seroconverters in the study (18.8%), yetour results may be less generalizable to this subpopulation.
The prospective nature of the study allowed the clinical ob-
servation of women after seroconversion. Overall, we observed11 cases of AIDS in the 54 women with subtype diagnosis. These women were followed for 199 person-years. The HIV-1subtype-specific AIDS incidence rates per 100 person-years var-ied widely, from 3.45 (95% confidence interval [CI], 1.44–8.29)for subtype A to 15.95 for subtype C (95% CI, 5.14–49.45) and14.45 for subtype D (95% CI, 4.66–44.81). No cases of AIDSwere seen in the 6 women who were infected with subtype Grelatively late in the study period.
Kaplan-Meier disease-free survival analysis was performed
on seroincident women grouped by HIV subtype (figure 3A). The AIDS-free survival curves differed by HIV subtype (logrank test, P ϭ .06; Wilcoxon-Gehan, P ϭ .03). At 5 years ofinfection, 87.0% of women infected with subtype A were AIDS-free compared with 30.0% of subtype C– and 60.0% of subtypeD–infected women. To adjust for other potentially confoundingvariables, we constructed a Cox proportional hazards modelwith AIDS as the outcome, controlling for nationality, age atregistration, years of registered prostitution, and HIV subtype. Figure 2.
HIV-1 subtypes in 54 registered female sex workers ob-
The adjusted ratio for development of AIDS was 8.11 for sub-
served from 1985 to 1997 with first seronegative visit (⅜), seroconver-sion date, subtype (A, C, D, G, and A/G), and AIDS status (᭢) in-
type C (95% CI, 1.44–45.61; P ϭ .02) and 8.43 for subtype D
dicated. Horizontal line to left of seroconversion date/subtype indicates
negative person-years of observation. Horizontal line to right of sero-
Since relatively few women were infected with subtypes C,
conversion date indicates positive person-years of observation. Cal-
D, and G, we analyzed AIDS-free survival probabilities in
women with subtype A compared with those with non-subtypeA HIV (figure 3B). The AIDS-free survival curves were distinct:
AIDS than those infected with subtype A (95% CI, 1.70–39.8;
Women infected with subtype A had a longer AIDS-free sur-
vival period than those infected with non-A subtypes; the 5-
In the prospective cohort study design, the evaluation of loss
year AIDS-free survival probability was 87% in subtype A–
to follow-up is critical to the interpretation of the AIDS pro-
infected women versus 52% for non-subtype A–infected women
gression data. At the beginning of each calendar year, study
(log rank test, P ϭ .02; Wilcoxon-Gehan, P ϭ .03). The Cox
physicians and social workers actively investigated enrolled
proportional hazards model estimated that women infected
women who had not attended the clinic for 112 months. In the
with a non-A subtype were 8.23 times more likely to develop
54 HIV-incident women with known HIV subtype, 4 (7.4%)
Figure 3. A, Kaplan-Meier AIDS-free survival in seroincident women by HIV-1 subtype (P ϭ .06, log rank; P ϭ .03, Wilcoxon-Gehan). B,
Kaplan-Meier AIDS-free survival analysis in seroincident women with HIV subtype A vs. all non-A subtypes (P ϭ .02, log rank; P ϭ .03, Wilcoxon-Gehan).
were considered lost to follow-up after the 1997 active follow-
global HIV pandemic. Differences in sexual practices, rates of
up evaluation. We performed a sensitivity analysis in which
STDs, and other determinants of HIV infection and transmis-
these women were considered to have developed AIDS. Again,
sion also likely contribute to the different HIV epidemic curves
subtype A demonstrated a significantly longer AIDS-free sur-
observed worldwide. However, to date, studies of different sub-
vival and reduced rate ratio for AIDS development compared
types have been limited to cross-sectional surveys. This is the
with non-A subtypes (95% CI, 0.07–0.74; P ϭ .014). Thus, we
first study that documents the introduction and spread of mul-
do not believe that differential follow-up in our study popu-
tiple HIV subtypes in a high-risk population studied over time.
lation was responsible for the subtype differences in progression
Previous studies have estimated the rate of progression to
AIDS in US and European cohorts. Although study designs
Misclassification bias must also be considered in the evalu-
and methods of analysis have varied, 65%–80% AIDS-free sur-
ation of our study results. Multiple methods have been used
vival at 5 years has been reported in cohorts before the extensive
to assess viral subtype infection, including heteroduplex mo-
use of antiviral therapy [20–23]. The rate of disease progression
bility shift assays, complete sequencing of the viral genome,
in populations in the developing world has been poorly studied
and PCR sequencing of partial regions of the HIV genome [2,
to date; our study cohort in Dakar, Senegal, provides the only
3, 18]. Sequence analysis of the V3 envelope region is one of
data on AIDS progression in an incident cohort studied 15
the more common methods for molecular epidemiology studies
years [8]. Studies concerning the natural history of chronic in-
and has relatively high reliability in determining HIV-1 subtypes
fections, such as HIV, are difficult to achieve. Nevertheless, our
[2]. However, this does not rule out the possibility of mosaic
study has been successful in minimizing loss to follow-up
genomes, which might only be detected through full genomesequencing [15–17]. Current data suggest that the number and
through regular active follow-up and the efforts of a dedicated
complexity of HIV recombinants is ever expanding. The ma-
jority of sequences from subtype G viruses originate in West
We believe our study is the first to evaluate disease progres-
and Central Africa; our data suggest this may be a more recent
sion in non-B subtype infections with known time of infection.
HIV-1 subtype [2]. In addition, sequence analysis of subtype G
In evaluating AIDS-free survival curves of women with incident
viruses suggest that this subtype may frequently be a mosaic
A, C, D, and G infection, we found distinct differences in AIDS-
with subtype A [19]. In consideration of this, we grouped sub-
free survival. The grouped comparison of non-A versus A sub-
types A and G together and compared disease progression rates
types demonstrated a significantly longer AIDS-free survival
to those of subtypes C and D. Subtypes A and G demonstrated
for women infected with subtype A. Due to the small sample
a significantly longer AIDS-free survival and adjusted AIDS
size per subtype, with few AIDS cases despite a lengthy study
rate ratio (95% CI, 1.70–39.82; P ϭ .009). Therefore, despite
period, our estimate of AIDS incidence should not be consid-
the possibility of mosaic viruses, our classification of subtypes
ered precise, and further study is clearly warranted. Cross-sec-
based on potential mosaicism still suggests distinct differences
tional studies indicate a significant proportion of AIDS cases
with subtype A infection in West and East Africa (unpublisheddata) [24]. However, this is not inconsistent with our results
Discussion
should the proportion of subtype A in the high-risk population
In recent years, there has been a growing appreciation of the
be quite elevated. Further study of HIV-1 subtype natural his-
role of HIV-1 subtypes in the complex epidemiology of the
tory and progression from different geographic regions is clearly
needed to better evaluate the role of viral subtype differences
7. Donnelly C, Leisening W, Sandberg S, Kanki P, Awerbach T. Comparison
of transmission rates of HIV-1 and HIV-2 in a cohort of prostitutes in
Senegal. Bull Math Biol 1993; 55:731–41.
The recognition of the different HIV-1 subtypes has led many
8. Marlink R, Kanki P, Thior I, et al. Reduced rate of disease development
to question whether properties of HIV-1 infection and its con-
with HIV-2 compared to HIV-1. Science 1994; 265:1587–90.
sequences as a whole can be generalized among different sub-
9. Weniger BG, Takebe Y, Ou CY, Yamazaki S. The molecular epidemiology
types. Our study provides the first evidence that HIV-1 subtypes
of HIV in Asia. AIDS 1994; 8:S1.
may differ in their progression time to AIDS. The further in-
10. Yu XF, Wang Z, Beyrer C, et al. Phenotypic and genotypic characteristics
of human immunodeficiency virus type 1 from patients with AIDS in
vestigation of these differences may be central to understanding
northern Thailand. J Virol 1995; 69:4649–55.
the diversity of HIV-1 pathogenesis. The evolution of HIV-1
11. Kanki P, Mboup S, Marlink R, et al. Prevalence and risk determinants of
subtypes in human populations may have favored or even se-
human immunodeficiency virus type 2 (HIV-2) and human immunode-
lected for diverse biologic properties. Such diversity could po-
ficiency virus type 1 (HIV-1) in West African female prostitutes. Am J
tentially affect the overall efficacy of HIV therapy or interven-
Epidemiol 1992; 136:895–907.
12. CDC. 1993 revised classification system for HIV infection and expanded
tion strategies. Thus, the study of the natural biologic variation
surveillance case definition for AIDS among adolescents and adults.
of the virus itself may provide important insights into the ra-
MMWR Morb Mortal Wkly Rep 1992; 41:1–19.
tional design of interventions to alter or interrupt disease
13. Wolinsky S, Wike CM, Korber BT. Selective transmission of human im-
munodeficiency virus type-1 variants from mothers to infants. Science
1992; 255:1134–7. Acknowledgments
14. Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sen-
sitivity of progressive multiple sequence alignment through sequence
weighting, position-specific gap penalties and weight matrix choice. Nu-
We thank Jim Donahue, Geoffrey Eisen, Brian Foley, David Hunter,
cleic Acids Res 1994; 22:4673–80.
John McNeil, and Boris Renjifo for critical review of this manuscript,
15. Siepel AC, Halpern AL, Macken C, Korber BTM. A computer program
Rajiv Rawat for editorial assistance, and the Senegalese Ministry of
designed to rapidly screen for HIV-1 intersubtype recombinant sequences.
Health for its support of this research.
AIDS Res Hum Retroviruses 1995; 11:1413–25.
16. Carr JK, Salminen MO, Gotte D, Koch C, Burke DS, McCutchan FE.
Mosaic structure of the full-length genome from a human immunodefi-
References
ciency virus type 1 isolate of clade E from Thailand. J Virol 1996; 70:
1. Over M, Piot P. Human immunodeficiency virus infection and other sexually
17. Gao F, Robertson DL, Morrison SG, et al. The heterosexual HIV-1 epidemic
transmitted diseases in developing countries: public health importance and
in Thailand is caused by an intersubtype (A/E) recombinant of African
priorities for resource allocation. J Infect Dis 1996; 174(suppl):S162–75.
origin. J Virol 1996; 70:7013–29.
2. Myers G, Korber B, Hahn B. Human retroviruses and AIDS 1995: a com-
18. Delwart EL, Shpaer EG, Louwagie J, et al. Genetic relationships determined
pilation and analysis of nucleic acid and amino acid sequences. Los Al-
by a DNA heteroduplex mobility assay: analysis of HIV-1 env genes.
amos, NM: Los Alamos National Laboratory, Theoretical Biology and
Science 1993; 262:1257–61.
Biophysics Group, 1995.
19. McCutchan F, Salimen MO, Carr JK, Burke DS. HIV-1 genetic diversity.
3. Louwagie J, McCutchan F, Peeters M. Phylogenetic analysis of gag genes
AIDS 1996; 10:S13–20.
from 70 international HIV-1 isolates provides evidence for multiple gen-
20. Lifson AR, Rutherford GW, Jaffe HW. The natural history of human im-
otypes. AIDS 1993; 7:769–80.
munodeficiency virus infection. J Infect Dis 1988; 158:1360–7.
4. Burke DS, McCutchan FE. Global distribution of human immunodeficiency
21. Moss AR, Bacchetti P. Natural history of HIV infection. AIDS 1989; 3:55–61.
virus-1 clades. In: Vincent T, DeVita J, Hellman S, Rosenberg SA, eds.
22. Alcabes P, Munoz A, Vlahov D, Friedland GH. Incubation period of human
AIDS: biology, diagnosis, treatment and prevention. Philadelphia: Lip-
immunodeficiency virus. Ann Epidemiol 1993; 15:303–18.
pincott-Raven, 1997:119–26.
23. Biggar RJ. The International Registry of Seroconverters. AIDS incubation
5. Leitner T, Alaeus A, Marquina S, Lilja E, Lidman K, Albert J. Yet another
in 1891 HIV seroconverters from different exposure groups. AIDS 1990;4:
subtype of HIV type 1? AIDS Res Hum Retroviruses 1995; 11:995–7.
6. Kanki P, Travers K, Hernandez-Avila M, et al. Slower heterosexual spread
24. Janssens W, Buve A, Nkengasong J. The puzzle of HIV-1 subtypes in Africa.
of HIV-2 compared with HIV-1. Lancet 1994; 343:943–6.
AIDS 1997; 11:705–12.
Stigmatisation vue de l’intérieur Témoignage de Lucie Boissinot dans le cadre de la journée de sensibilisation à la détection et au traitement précoce des premières psychoses. ______________________________________________________________ Le pire des maux en ce qui me concerne, c’est la maladie et non la Lorsque mon fils est tombé malade, il m’a été très difficile d
Consumer Medication Brochure Series What is an antipsychotic? T h e m a i n a d v a n t a g e o f t h e a t y p i c a l How should I take the medication? An antipsychotic is a medication that is used to antipsychotics is that they are less likely to cause You should take the medication as prescribed by help treat psychotic symptoms. Psychotic side effects such as muscle st
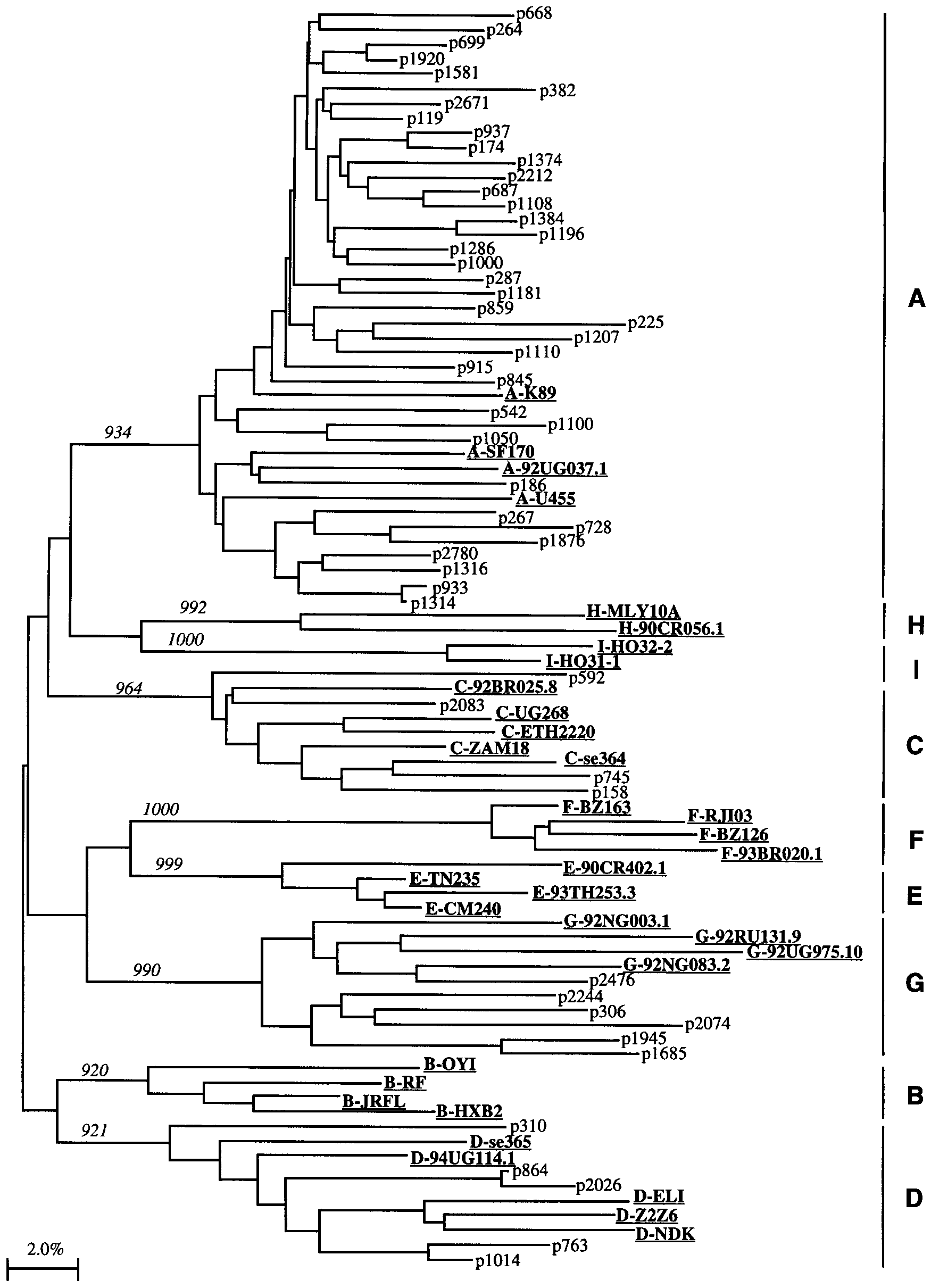 Figure 1.
Figure 1.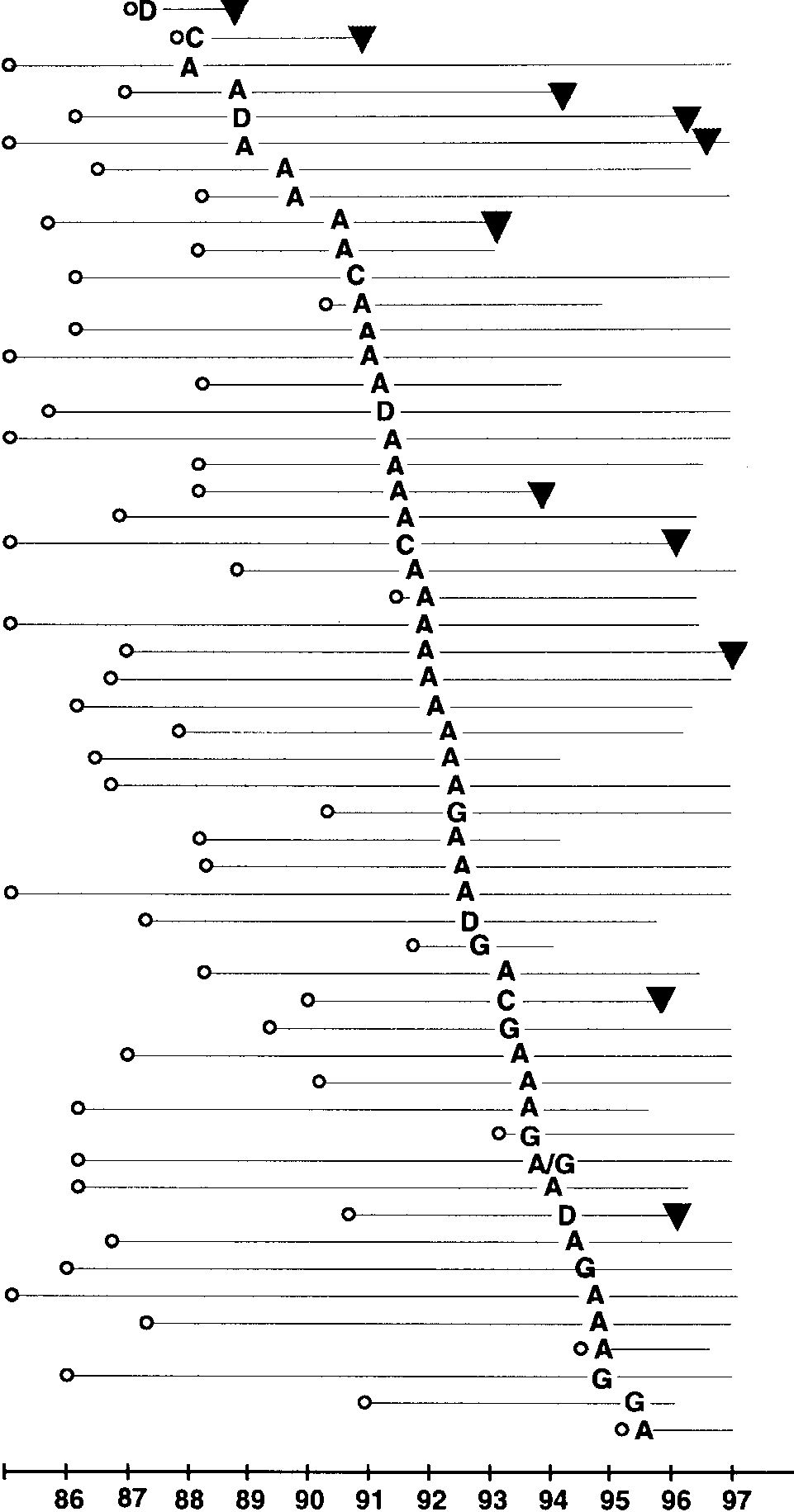 illustrated by HIV subtype designated at the estimated sero-conversion date (figure 2). In this high-risk group of women,the first HIV-1 seroconversions occurred in late 1987. Through-out the study period, subtype A was the most common HIV-1 subtype. The first seroconversion to subtype C occurred latein 1987; the last C seroconversion was observed in late 1993.
illustrated by HIV subtype designated at the estimated sero-conversion date (figure 2). In this high-risk group of women,the first HIV-1 seroconversions occurred in late 1987. Through-out the study period, subtype A was the most common HIV-1 subtype. The first seroconversion to subtype C occurred latein 1987; the last C seroconversion was observed in late 1993.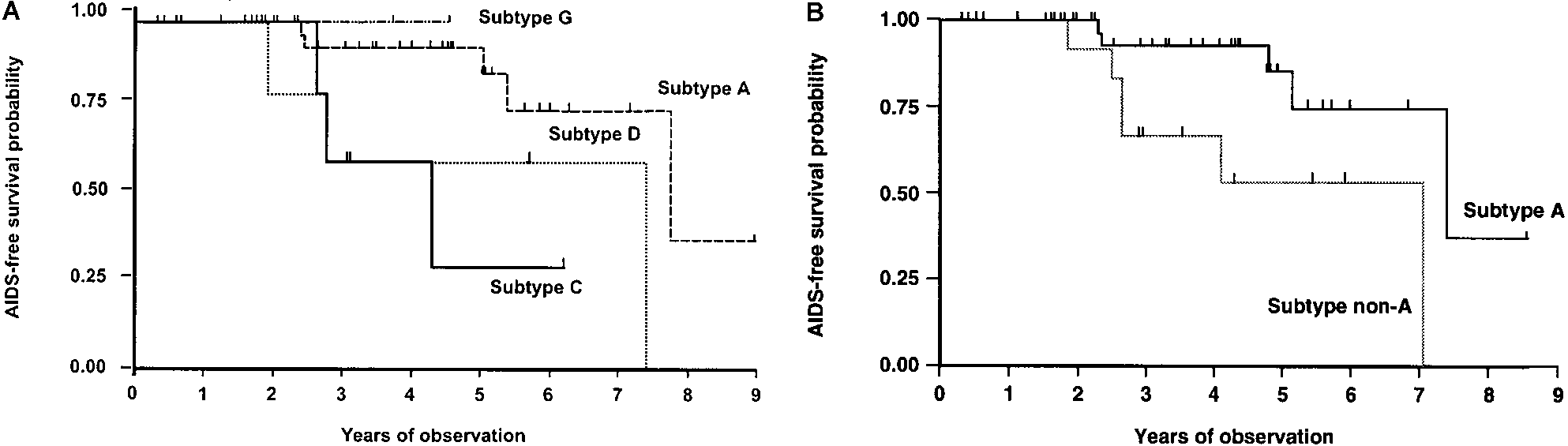 Figure 3.
Figure 3.