Levitra has a minimal amount of contraindications which has increased its popularity kamagra uk You can buy quality certified medications from us at an affordable price.
S. 355-359, 168/97
Clin Chem Lab Med 1998; 36(6):355–359 1998 by Walter de Gruyter · Berlin · New York
Engineering of an Anti-Steroid Antibody: Amino Acid Substitutions Change Antibody Fine Specificity from Cortisol to Estradiol Patrick Chames and Daniel Baty
be coupled to immunogenic proteins. The specificity ofan anti-steroid antibody depends on the position of
Laboratoire d’Ingénierie des Systèmes Macromoléculaires,Marseille, France
coupling to the carrier protein (6). The ring coupling po-sition determines the cross-reactivity of the antibody(7). Cross-reactivity with different steroids may result
Immunoassays are widely used for determination of
from small but significant conformational changes in
the concentration of steroid hormones. However, ob-
the antibody which allow it to accommodate different li-
taining specific anti-steroid monoclonal antibodies re-
gand orientations in the binding site (8). For all these
mains difficult. We used antibody engineering and
reasons, no anti-estradiol antibody able to detect pico-
phage display methods to change the specificity of an
grammes of estradiol in plasma and with no cross-reac-
anti-cortisol monoclonal antibody towards estradiol.
tion against related steroids, has ever been produced. This work demonstrates that production of recombi-
Remodelling the antibody-combining site could be
nant antibodies may be a valuable way of obtaining
used to increase the specificity of an antibody. Genetic
the high-specificity antibodies required for steroid im-
engineering techniques may make it possible to im-
munoassays.
prove the binding properties of potentially useful anti-
Key words: Antibody; Cross-reactivity; Phage display;
bodies by using mutagenesis to modify the amino acid
Single-chain variable fragment (scFv); Steroid.
sequence within the antibody-combining site. Substi-tutions in only a few amino acids can increase affinity(9, 10), decrease cross-reactivity with closely-related
Introduction
antigens (9) or change specificity (11). The use of phagedisplay technology has made possible the genetic en-
Steroid hormones have a number of different effects
gineering of immunoglobulins. It is possible to mimic
on their target tissues. They are involved in many cellu-
the immune system in vitro by expressing antibody
lar processes controlling metabolism, reproduction,
fragments on the surface of filamentous bacterio-
differentiation and behaviour (1). Many clinical symp-
phages (12). Antibody fragments expressed at the
toms are associated with abnormal steroid levels. Syn-
phage surface can be selected on the basis of specific-
thetic steroids are widely used as agonists or antago-
ity, affinity, avidity, or binding kinetics. Genes coding
for antibody fragments can be mutated by PCR meth-
Steroids are small molecules (300–400 Da) biosyn-
ods to increase affinity (13, 14), change specificity (15)
thesized from cholesterol. Their molecular structures
or modify cross-reactivity (16, 17). Short et al. obtained
are very similar. Estradiol has a hydroxyl group at C17,
two anti-digoxin Fab mutants with affinities two and
whereas estrone has a ketone group. Cortisol is pro-
four times higher than that of the parental antibody
duced from its precursor, 11-deoxycortisol, by hydroxy-
from a phage display library containing randomized
lation at C11. Measurement of steroid concentration is
mutations of heavy chain residues (18).
very useful during therapy and for clinical diagnosis. It
X-ray crystallographic data from antibody fragments
is difficult to measure hormone concentrations in target
indicate that only a few residues of the antibody-com-
cells, but blood hormone concentrations and hormone
bining site contribute significantly to affinity. The bind-
metabolites in urine can be determined. A competitive
ing site must have some plasticity, particularly when
binding assay, using specific antibodies with high affin-
the antigens themselves are rigid, as is the case for
ity and radioactive steroids, is the most commonly used
steroids (19). The cross-reactivity of an anti-progeste-
method for measuring steroid concentrations.
rone Fab for steroid analogues may be due to flexibility
However, the structural similarities of steroids often
of the antibody-combining site (20). Molecular model-
cause problems with cross-reaction in immunoassay.
ling of the antibody-combining site may be a valuable
Plasma and urine both contain substances that cross-
approach for identifying residues in contact with the
react with anti-steroid antibodies. The steroids can be
steroid where the X-ray structure is not known (21).
extracted with an organic solvent and separated by
We have described the production in Escherichia coli
chromatography before measuring their concentration
(E. coli) of an anti-cortisol single-chain variable frag-
by immunoassay (2–4). This method is laborious and
ment (scFv 5A4; where the variable domains VH and VL
direct immunoassay on plasma or urine extracts is pref-
are covalently associated by a flexible peptide) (22).
erable for multiple determinations. Unfortunately, the
The corresponding 5A4 monoclonal antibody (mAb)
method used for immunoassay can significantly affect
recognizes cortisol with a dissociation constant of 10-8
the results obtained (5). Moreover, steroids cannot
M (22). Like most anti-steroid antibodies, this mAb
themselves cause an immunological reaction and must
cross-reacts with steroid analogues but does not rec-
Chames and Baty: Engineering of an anti-steroid antibody
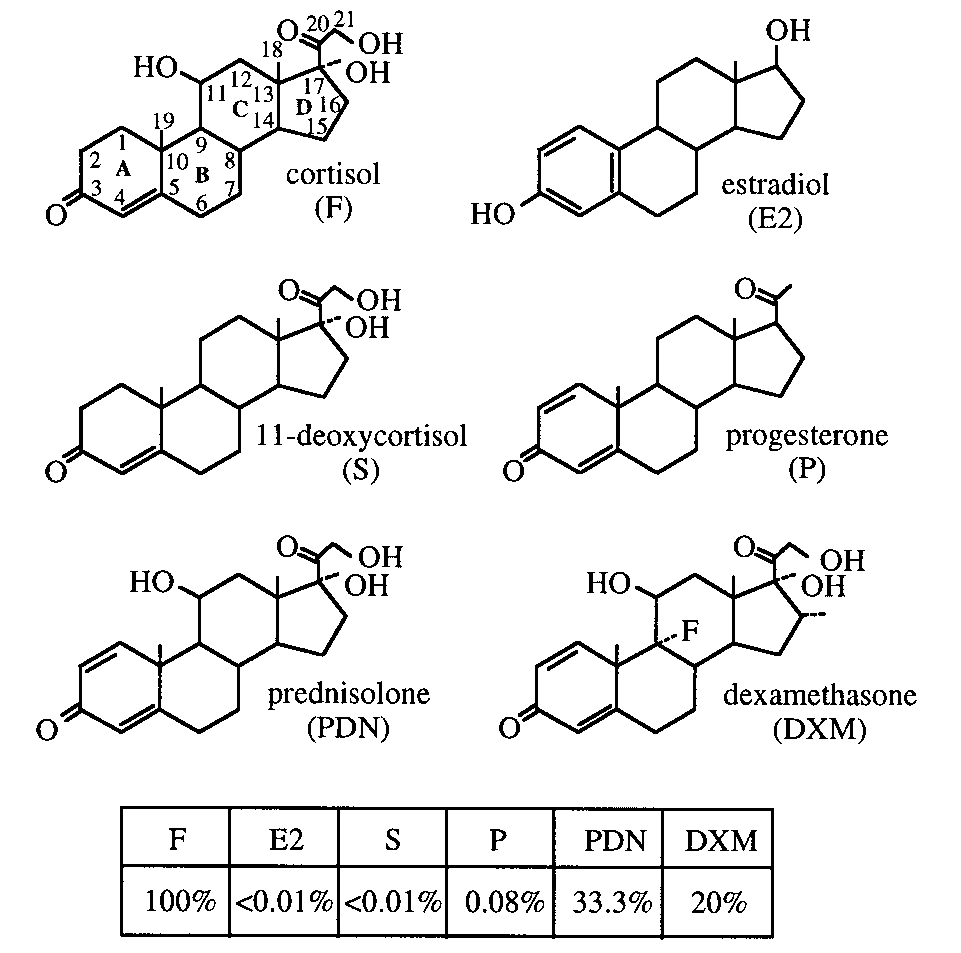
ognize estradiol (Fig. 1). We have also described the
Materials and Methods
identification of the residues of the antibody-combin-ing site and those which are likely to interact with corti-
sol (22). In the present study, we used mutagenesis to
The E. coli strain TG1 was used as the bacterial host for the
change the specificity of this anti-cortisol antibody into
preparation of phagemids, and as the host of bacteriophage
Construction of the wild-type anti-cortisol scFv
To construct the 5A4 scFv, VH and VL genes were amplifiedfrom the plasmid pscFv5A4 by PCR (22). The 3’ primer used toamplify VH contained part of the coding sequence for thelinker (Gly3Ser)4 and the 5’ primer used to amplify VL con-tained the rest of the sequence (with 15 overlapping bases). The scFv was assembled by splice overlap extension PCR (23). At the same time, two restriction enzyme sites (NcoI and EagI)were introduced at the 5’ and 3’ ends, respectively, of the scFvgene to facilitate cloning into the pHENI phagemid (24) to givepHENscFv5A4.
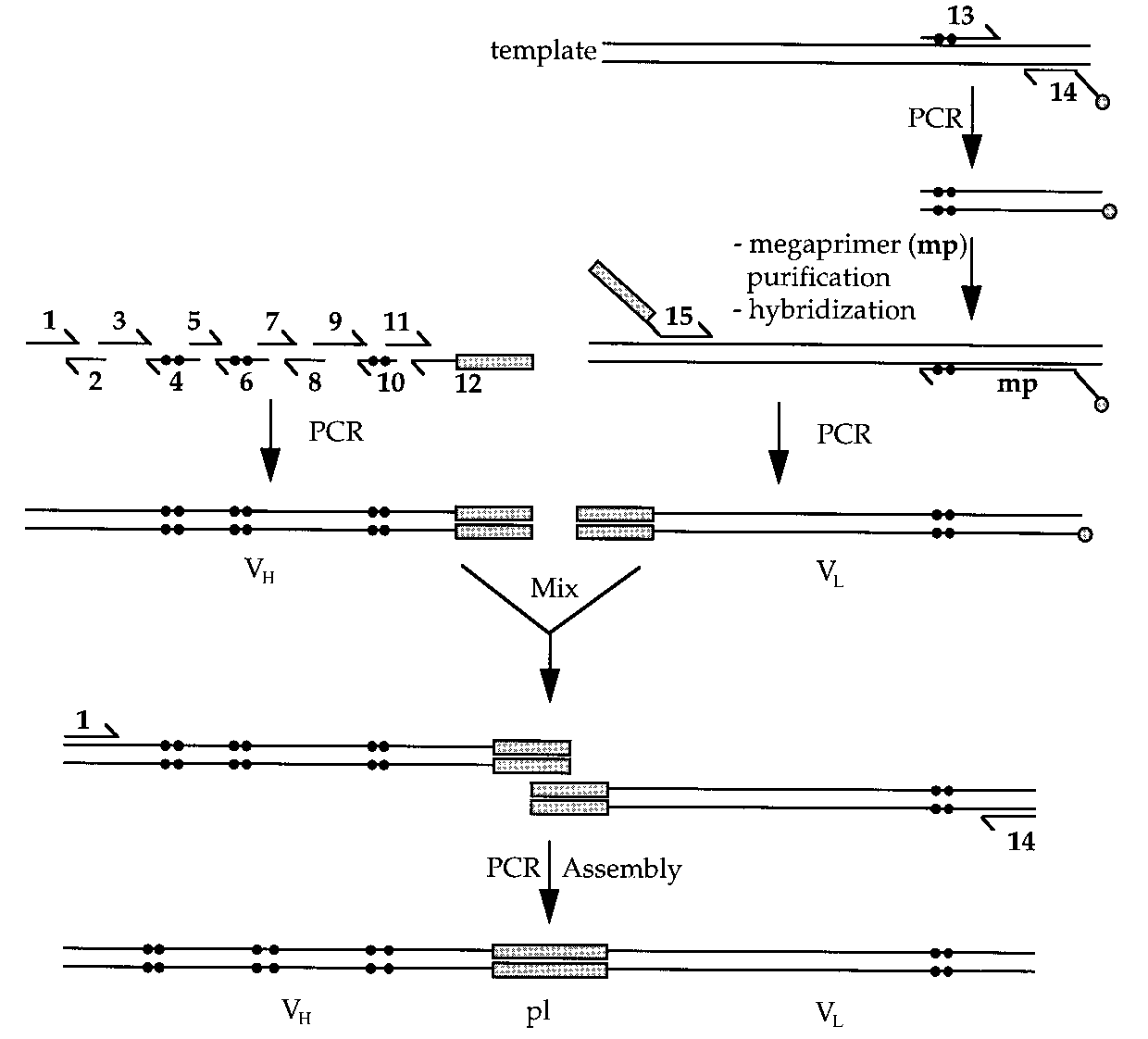
We constructed the mutant VH library using recursive PCRmethods (25) and a set of overlapping oligonucleotides (Fig. 2). The mutant VL library was constructed with a new PCR-based site-directed mutagenesis method using magneticbeads (26).
The oligonucleotides used for PCR are: oligo 1: 5’ CTCGC
KGCSCAGCCGGCCATGGC 3’; oligo 2: 5’ CCCAGGCTTCACC
Structures of cortisol (F), estradiol (E2), 11-deoxycorti-
AGCTCAGTTCCTGACTGCTGCAGCTGGACCTCGGCCATGGC
sol (S), progesterone (P), prednisolone (PDN), and dexameth-
CGGCTG 3’; oligo 3: 5’ GAGCTGGTGAAGCCTGGGGCTTCA
asone (DXM) and their cross-reactivity with monoclonal anti-
GTGAAGCTGTCCTGCAAGACTTCTGGCTTCACCTTCAGCAG
body 5A4. The cross-reactivities determined by equilibrium
TAGG 3’; oligo 4: 5’ CCAGACTTTTGCTTCAACCAVNNCATVN
dialysis are given as the percentage of cortisol concentration
NCCTACTGCTGAAGGTG 3’; oligo 5: 5’ GGTTGAAGCAAAAGT
divided by steroid concentration at 50% binding of the zero
CTGGACAGAGTCTTGAGTGGATTGC 3’; oligo 6: 5’ GCCTGTG
PCR assembly of the scFv library. Oligonucleotides
(mp) was mixed with the template and oligonucleotide 15 for a
1–12 were mixed and subjected to PCR amplification to create
second PCR amplification. The two mutated PCR fragment li-
the mutated VH library. The mutated VL library was constructed
braries (VH and VL) were mixed with oligonucleotides 1 and 14
using the wild-type phagemid template pHENscFv5A4 and oli-
and amplified by PCR to give the mutated scFv library. The dots
gonucleotides 13 and 14 for the first PCR amplification. After
mark the mutated positions. The boxes represent the coding
purification, the biotinylated single stranded megaprimer
sequence for the peptide linker (pl).
Chames and Baty: Engineering of an anti-steroid antibody
A A C T T C T G AT TATA V N N A G A A C C A C C A G T T C C A C C A
Results and Discussion
T AAATVNNTGCAATCCACTCAAGACTC 3’; oligo 7: 5’ AATCAGAAGTTCACAGGCAAGGCCCAAGTGACTGTAGACACATC 3’;
We used the 5A4 antibody to create a library of anti-
oligo 8: 5’ GCTGAATTGCATGTAGACTGTGCTGGAGGATGTG
steroid antibody fragments by site-directed mutagene-
TCTACAGTCAC 3’; oligo 9: 5’ CTACATGCAATTCAGCAGCCT-
sis. The objective was to use phage display to select
GACAACTGAGGACTCTGCCATCTATTACTGTGCAAG 3’; oligo
from the mutants the antibody fragment with the re-
10: 5’ GGCCCCAGGAATCAAAGGGVNNGCTGCTACCATAG quired specificity.
ACACTVNNTCTTGCACAGTAATAGATGG 3’; oligo 11: 5’ TTGATTCCTGGGGCCAAGGCACCTCAGTCACCGTCTCCTCA 3’;
We have previously cloned the scFv fragment (heavy
oligo 12: 5’ TCACCTGAACCAGGTTTACCAGAACCTGAGGTA-
and light variable domains linked by a flexible peptide
GAACCTGAGGAGACGGGTGAC 3’; oligo 13: 5’ GCCAGCAGT
linker) of the 5A4 mAb and sequenced its gene (22). We
expressed scFv fused to the g3p protein at the M13
AGBTGGA 3’; oligo 14: 5’ CAAGCTTACTAGTTTATGCGGCCC-
phage surface, and used phage ELISA experiments to
CATTCAGATCC 3’; oligo 15: 5’ CTGGTAAACCTGGTTCAGGT
show that scFv had the same pattern of cross-reaction.
GAAGGTAGTACTAAAGGTGACATTGTGCTGAC 3’. The mu-
The phage antibody immunoassay is a highly sensitive
tated positions are indicated by italics (B = G, T or C; K = G or
detection system. It facilitates the characterization of
T; N = A, C, G or T; S = G or C; and V = G, A or C). The codon NNB
antibody fragments and eliminates the need for purifi-
was used to reduce the number of stop codons. The PCR prod-
cation of antibodies (30). The 5A4 phage antibody
ucts were purified by electrophoresis in polyacrylamide gelsand the two libraries were assembled by splice overlap exten-
(Phab) had very similar cross-reactivity to the parental
sion. NcoI and EagI sites (underlined) were introduced at the
monoclonal antibody, as is often the case (31). We have
same time. The final, purified PCR product (5 µg) was inserted
previously built a molecular model of the 5A4 paratope
into the pHENI phagemid (2 µg). Fifty electroporations were
complexed with cortisol from the primary sequence
used to produce a library of 108 clones. Some of these clones
and information about recognition of cortisol deriva-
were tested by miniprep and restriction analysis for the pres-
tives and analogues (22). This model allowed us to
ence of an insert of expected size. These clones were then
identify amino acids putatively in contact with the ster-
tested for expression of a 30 kDa product as previously de-
oid. We found that the steroid nucleus may be stacked
scribed (27). The library was rescued as previously described
between the side chains of two tryptophans. We de-
cided to conserve these two amino acids that could
interact with any steroid, and all amino acids close tothe steroid were randomized. Eight different positions
The antigen, estradiol-6-O-carboxymethyloxime-bovine
were chosen in four complementarity-determining re-
serum albumin (E2-6-CMO-BSA; 10 µg/ml in phosphate buf-
gions (CDR) (Tab. 1). The creation of a large library re-
fered saline (PBS) solution) was incubated in maxisorp immu-
quired large quantities of insert coding for scFv. These
notubes (Nalge Nunc International, Naperville, IL, USA) over-night at 4 °C. The tubes were then saturated with 2 % milk PBS
mutations were mapped to sites along the entire length
(MPBS) and incubated for one hour at 37 °C. We used 1013
of the gene. Therefore, we used recursive PCR methods
titered units of library phages in a total volume of 2 ml MPBS
(25) to introduce six mutations in the VH gene and a new
per immunotube for the first round of selection. Immunotubes
PCR-based method using magnetic beads to improve
were washed and bound phages were eluted as previously
the mutagenesis efficiency in VL (26). Splice overlap ex-
described (28). After each round of selection, E. coli TG1 were
tension (23) was then used to assemble the VH and VL li-
reinfected with eluted phages and rescued to provide phages
brary (Fig. 2). The DNA was purified, digested, ligated,
for the next round of selection. Selection was stopped when
and electroporated to produce a library of 108 different
the recovery of eluted phages increased significantly (>10
clones. Library quality was tested by DNA restriction
analysis and Western blot, and 96 % of clones contained
Screening of clones using enzyme-linked immunosorbent as-
a full length insert and 36 % of clones expressed scFv
detected by Western blotting using IPTG induction andthe 9E10 antibody to detect the c-myc tag. Vaughan et
Single ampicillin-resistant colonies, resulting from infection
al. obtained similar results from their library (32). The
of E. coli TG1 with eluted phages were used to inoculate 150 µl
low percentage of scFv-expressing clones may be due
of 2YT medium containing ampicillin 100 µg/ml and glucose 2 %, in 96-well plates. Overnight production of phages was as
to frame-shifts, deletions or creation of stop codons in-
previously described (28). Supernatants containing phages
troduced during the three successive PCR steps.
were tested for binding by enzyme-linked immunosorbent as-
Residues forming the antibody-combining site in the
say (ELISA) in Falcon 96-well plates coated with antigens (in-
5A4 scFv model. The mutated residues of the library are in
cubated at 10 µg/ml in PBS overnight) and saturated with
MPBS. Binding of phages was detected with a horseradishperoxidase-anti-M13 antibody conjugate (Amersham Phar-
Analysis of phage binding to steroid-BSA was performed us-
ing bacterial supernatants containing phage, essentially as
H95, T100B, F100D
previously described (29, 30), with 10 µg/ml antigen in PBS in
all cases, in Falcon 96-well plates.
W91, N94, P96
Chames and Baty: Engineering of an anti-steroid antibody
The 5A4 Phab was not cross-reactive with estradiol,
a steroid very different in structure to cortisol. So, afterrescue using a phage helper, the library was selectedagainst estradiol-6-CMO-BSA. Positive Phabs were de-tected by ELISA after the third round of selection, andalmost all Phabs eluted after the fourth round werepositive (Tab. 2). Sequencing of 18 positive Phabsshowed that a single clone had been selected. The VHsequence of this clone (named 1B8) was identical tothat of the wild-type scFv. However, the VL gene hadtwo mutations (Asn94Arg and Pro96Asp) in the CDR3. The 1B8 Phab was further characterized by ELISA (Fig. 3). Cross-reaction of the 5A4 Phab with prednisolonewas not detectable with the 1B8 Phab. The 1B8 Phabgave a strong signal with estradiol but had residualbinding to cortisol. Estradiol has a hydroxyl group atposition C3. This position is close to the two mutationsin the mutant Phab in our model. These new amino ac-ids may form a hydrogen bond at this position, allow-ing recognition of estradiol. However, only part of the
Specificity of 5A4 and 1B8 Phabs. ELISA was used to
original paratope was modified and this may explain
test Phabs for their ability to bind directly to steroid-BSA. Corti-
the residual binding to cortisol. It is highly improbable
sol-3-O-carboxymethyloxime-BSA (F-3-CMO-BSA), cortisol-21-
that selection led to conservation of wild-type codons
hemisuccinate-BSA (F-21-HS-BSA), 11-deoxycortisol-3-CMO-
in the VH gene. The selected wild-type VH gene may re-
BSA (S), prednisolone-3-CMO-BSA (PDN), progesterone-3-
sult from contamination during the construction of the
CMO-BSA (P), estradiol-6-CMO-BSA (E2), BSA or milk (T) were
mutant library. It may also indicate that the gene cod-
used to coat 96-well plates. Culture supernatants containing
Phabs were incubated with the plates for one hour at room tem-
H domain in the original library may have
been mutated too much, destabilizing the paratope
perature. The plates were washed and bound Phabs were de-
scaffold. This problem could be overcome by using the
tected by addition of horseradish peroxidase/anti-M13 conju-gate. The absorbance, indicated by bars, was measured at
parsimonious mutagenesis method that limits the
number of amino acids that do not retain parentalstructural features (33). The mutation rate is controlledand can be limited to two or three mutations per clone
Acknowledgements
over the eight degenerated positions.
We thank C. Y. Cuilleron and E. Mappus for the gift of steroid-
Frequency of binding clones from the scFv-phage li-
BSA and helpful discussion. We are very grateful to D. Duché
brary. The number of phage-transfecting units (T. U.) is given
and J. Chauveau for critical reading and helpful discussion
before (input) and after (output) each round of selection. Forty
and to M. Chartier for technical assistance. This work was sup-
eight clones in each round of selection were tested by ELISA
ported by the Centre National de la Recherche Scientifique.
for their capacity to bind estradiol-6-CMO-BSA. References
1. Milgrom E. Steroid hormones. In: Baulieu EE, Kelly PA, ed-
itors. Hormones: from molecules to disease. Paris: Her-
2. Barkley MS, Lasley BL, Thompson MA, Shackleton CH.
Equol: a contributor to enigmatic immunoassay measure-ments of estrogen. Steroids 1985; 46:587–608.
We selected a new anti-estradiol antibody fragment
3. Agasan AL, Stewart BJ, Watson TG. Development of a ra-
by molecular modelling, site-directed randomization
dioimmunoassay method for ethynylestradiol in plasma
and phage display. The specificity of this clone could be
using a monoclonal antibody. J Immunol Methods 1994;
improved. For example, chain-shuffling (34) could be
4. Wudt SA, Wachter UA, Homoki J, Teller WM. 17 alphahy-
used to select a VH partner that specifically interacts
droxyprogesterone, 4-androstenedione, and testosterone
with another site of the steroid. The resulting clone
profiled by routine stable isotope dilution/gas chromato-
may have higher affinity for estradiol and may lose its
graphy-mass spectrometry in plasma of children. Pediatr
affinity for cortisol. This work demonstrates that anti-
body engineering is a valuable technique for obtaining
5. Cook NJ, Read GF. Oestradiol measurement in women on
specific anti-steroid antibodies for immunoassays.
oral hormone replacement therapy: the validity of com-mercial test kits. Br J Biomed Sci 1995; 52:97–101.
6. Yoon DY, Choi MJ, Choe IS, Chung TW, Byun SM. Influence
of the conjugation site on the specificity of monoclonal
Chames and Baty: Engineering of an anti-steroid antibody
antibodies to progesterone and on the performance of di-
bovine anti-testosterone monoclonal antibody. Protein
rect enzyme immunoassay. Biochem Mol Biol Int 1993;
22. Le Calvez H, Fieschi J, Green JM, Marchesi N, Chauveau J,
7. Gani M, Coley J, Piron J, Humphreys AS, Arevalo J, Wilson
Baty D. Paratope characterization by structural modelling
IA, et al. Monoclonal antibodies against progesterone: ef-
of two anti-cortisol single-chain variable fragments pro-
fect of steroid-carrier coupling position on antibody speci-
duced in E. coli. Mol Immunol 1995; 32:185–98.
ficity. J Steroid Biochem Mol Biol 1994; 48:277–82.
23. Horton RM, Hunt HD, Ho SN, Pullen JK, Pease LR. Engi-
8. Arevalo JH, Hassig CA, Stura EA, Sims MJ, Taussig MJ,
neering hybrid genes without the use of restriction en-
Wilson IA. Structural analysis specificity. Detailed compar-
zymes: gene splicing by overlap extension. Gene 1989;
ison of five Fab’-steroid complexes. J Mol Biol 1994;
24. Hoogenboom HR, Griffiths AD, Johnson KS, Chiswell DJ,
9. Roberts S, Cheetham JC, Rees AR. Generation of an anti-
Hudson P, Winter G. Multi-subunit proteins on the surface
body with enhanced affinity and specificity for its antigen
of filamentous phage: methodologies for displaying anti-
by protein engineering. Nature 1987; 328:731–4.
body (Fab) heavy and light chains. Nucleic Acids Res 1991;
10. Sharon J. Structural correlates of high antibody affinity:
three engineered amino acid substitutions can increase
25. Prodromou C, Pearl LH. Recursive PCR: a novel technique
the affinity of anti-p-azophenylarsonate antibody 200-fold.
for total gene synthesis. Protein Eng 1992; 5:827–9.
Proc Natl Acad Sci USA 1990; 87:4814–7.
26. Fieschi J, Niccoli P, Camilla C, Chames P, Chartier M, Baty
11. Kussie PH, Parhami-Seren B, Wysocki LJ, Margolies MN. A
D. Polymerase chain reaction-based site-directed muta-
single engineered amino acid substitution changes anti-
genesis using magnetic beads. Anal Biochem 1996;
body fine specificity J Immunol 1994; 152:146–52.
12. Marks JD, Hoogenboom HR, Griffiths AD, Winter G. Molec-
27. Harrison JL, Williams SC, Winter G, Nissim A. Screening
ular evolution of proteins on filamentous phage: mimick-
of phage antibody libraries. Methods Enzymol 1996;
ing the strategy of the immune system. J Biol Chem 1992;
28. Marks JD, Hoogenboom HR, Bonnert TP, McCafferty J,
13. Deng SJ, MacKenzie CR, Sodowska J, Michniewicz J,
Griffiths AD, Winter G. By-passing immunization: human
Young NM, Bundle DR, et al. Selection of antibody single-
antibodies from V-gene libraries displayed on phage. J
chain variable fragments with improved carbohydrate
binding by phage display. J Biol Chem 1994; 269:9533–8.
29. Hoogenboom HR, Winter G. By-passing immunisation: hu-
14. Schier R, McCall A, Adams GP, Marshall KW, Merritt H, Yim
man antibodies from synthetic repertoires of germline VH
M, et al. Isolation of picomolar affinity anti-c-erbB-2 single-
gene segments rearranged in vitro. J Mol Biol 1992;
chain Fv by molecular evolution of the complementary de-
termining regions in the center of the antibody binding
30. Li Y, Owen MR, Cockburn W, Kumagai I, Whitelam GC.
site. J Mol Biol 1996; 263:551–67.
Study of antibody-antigen interaction through site-di-
15. Casson LP, Manser T. Evaluation of loss and change of
rected mutagenesis of the VH region of a hybrid phage-
specificity resulting from random mutagenesis of anti-
antibody fragment. Protein Eng 1996; 9:1211–7.
body VH region. J Immunol 1995; 155:5647–54.
31. Skerra A. Bacterial expression of immunoglobulin frag-
16. Glaser SM, Yelton DE, Huse WD. Antibody engineering by
ments. Curr Opin Immunol 1993; 5:256–2.
codon-based mutagenesis in a filamentous phage vector
32. Vaughan TJ, Williams AJ, Pritchard K, Osbourn JK, Pope
system. J Immunol 1992; 149:3903–13.
AR, Earnshaw JC, et al. Human antibodies with sub-nano-
17. Barbas CF, Hu D, Dunlop N, Sawyer L, Cababa D, Hendry
molar affinities isolated from a large non-immunized
RM, et al. In vitro evolution of a neutralizing human anti-
phage display library. Nature Biotechnol 1996; 14:309–14.
body to human immunodeficiency virus type 1 to enhance
33. Schier R, Balint RF, McCall A, Apell G, Larrick JW, Marks
affinity and broaden strain cross-reactivity. Proc Natl Acad
JD. Identification of functional and structural amino acid
residues by parsimonious mutagenesis. Gene 1996;
18. Short MK, Jeffrey PD, Kwong RF, Margolies MN. Contribu-
tion of antibody heavy chain CDR1 to digoxin binding ana-
34. Marks JD, Griffiths AD, Malmqvist M, Clackson TP, Bye JM,
lyzed by random mutagenesis of phage-displayed Fab 26-
Winter G. By-passing immunization building affinity hu-
10. J Biol Chem 1995; 270:28541–50.
man antibodies by chain shuffling. Biotechnology 1992;
19. Arevalo JH, Stura, EA, Taussig MJ, Wilson IA. Three-di-
mensional structure of an anti-steroid Fab’ and progeste-rone-Fab’ complex. J Mol Biol 1993; 231:103–18.
Received 5 November 1997; accepted 17 April 1998
20. Arevalo JH, Taussig MJ, Wilson IA. Molecular basis of the
crossreactivity and the limits of antibody-antigen comple-
Corresponding author: Dr. D. Baty, UPR 9027, CNRS, IBSM, 31
mentarity. Nature 1993; 365:859–63.
chemin Joseph Aiguier, F-13402 Marseille Cedex 20, France
21. Jackson T, Morris BA, Martin ACR, Lewis DVF, Sanders PG.
Tel.: +33 4 91 16 41 17, Fax: +33 4 91 71 21 24
Molecular modelling and site-directed mutagenesis on a
Work in Progress Published monthly by Michele Crawford Work in Progress is an electronic newsletter intended to assist individuals seeking optimum well-being. Mail to ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~ Work in Progress May 2006 Volume I Issue III Feature Article: Part One: Questioning the Treatment of Anxiety ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~ Please feel free to forward
How Consultations by People Living with HIV Drive Change and Shape Policies, Programs and Normative Guidelines Christoforos Mallouris, Georgina Caswell, and Edwin J. Bernard People living with HIV are increasingly involved in shaping HIV responses and their collective expertise and competencies are informing the revision and/or development of normative guidelines and policy framewo
 Chames and Baty: Engineering of an anti-steroid antibody
ognize estradiol (Fig. 1). We have also described the
Materials and Methods
Chames and Baty: Engineering of an anti-steroid antibody
ognize estradiol (Fig. 1). We have also described the
Materials and Methods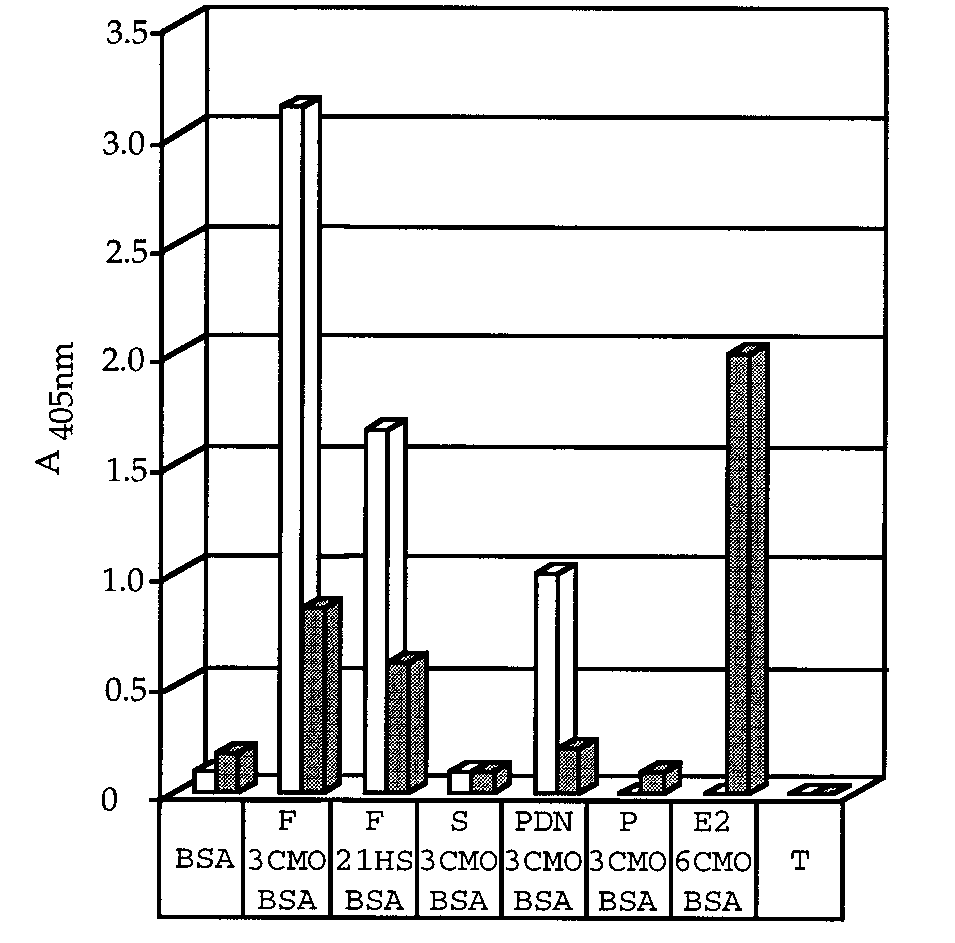 Chames and Baty: Engineering of an anti-steroid antibody
The 5A4 Phab was not cross-reactive with estradiol,
a steroid very different in structure to cortisol. So, afterrescue using a phage helper, the library was selectedagainst estradiol-6-CMO-BSA. Positive Phabs were de-tected by ELISA after the third round of selection, andalmost all Phabs eluted after the fourth round werepositive (Tab. 2). Sequencing of 18 positive Phabsshowed that a single clone had been selected. The VHsequence of this clone (named 1B8) was identical tothat of the wild-type scFv. However, the VL gene hadtwo mutations (Asn94Arg and Pro96Asp) in the CDR3.
Chames and Baty: Engineering of an anti-steroid antibody
The 5A4 Phab was not cross-reactive with estradiol,
a steroid very different in structure to cortisol. So, afterrescue using a phage helper, the library was selectedagainst estradiol-6-CMO-BSA. Positive Phabs were de-tected by ELISA after the third round of selection, andalmost all Phabs eluted after the fourth round werepositive (Tab. 2). Sequencing of 18 positive Phabsshowed that a single clone had been selected. The VHsequence of this clone (named 1B8) was identical tothat of the wild-type scFv. However, the VL gene hadtwo mutations (Asn94Arg and Pro96Asp) in the CDR3.