Levitra has a minimal amount of contraindications which has increased its popularity viagra uk You can buy quality certified medications from us at an affordable price.
Comparison of isoniazid oxidation catalyzed by bacterial catalase-peroxidases and horseradish peroxidase
Vol. 323, No. 2, November 10, pp. 438 – 446, 1995
Comparison of Isoniazid Oxidation Catalyzed by BacterialCatalase – Peroxidases and Horseradish Peroxidase1
Alex Hillar and Peter C. Loewen2Department of Microbiology, University of Manitoba, Winnipeg, Manitoba R3T 2N2, Canada
Received June 22, 1995, and in revised form August 31, 1995
Isonicotinic acid hydrazide (INH)3 is one of a number
The physical properties and activities of the purified
of drugs used to treat Mycobacterium tuberculosis in-
catalase – peroxidase hydroperoxidase I (HPI) of Esch-
fections. By contrast, INH is ineffective or has lower
erichia coli (EcHPI) and HPI with a carboxyl-terminal
efficacy when used against Mycobacteria other than
extension of Mycobacterium tuberculosis (MtHPI-e) M. tuberculosis or M. bovis and is ineffective against
are compared to those of commercial preparations of
strains of Escherichia coli even at high concentrations. horseradish peroxidase (HRP). The catalase – peroxi-
It has long been recognized that catalase-deficient
dase proteins had similar absorption spectra and dif-
strains of M. tuberculosis are resistant to treatment
fered primarily in that MtHPI-e has a higher peroxi-
with INH (1), but only recently has it been shown that
datic to catalatic activity ratio than EcHPI. Trypsin
either deletion (2) or point mutation (3) of the katGcleavage of MtHPI-e resulted in the formation of an
gene gives rise to such a phenotype. In M. tuberculosis,active catalase – peroxidase lacking the carboxyl-ter- katG encodes the only catalase, which also has an asso-
minal extension. The three enzymes, HRP, MtHPI-e,
ciated peroxidase activity. This catalase – peroxidase is
and EcHPI, mediated the isoniazid- and H2O2-depen-
50% identical to hydroperoxidase I of E. coli (EcHPI)
dent production of radical species, as detected by ni-
which in turn has been shown to be related to the su-
troblue tetrazolium reduction. A constant flux of H2O2,
perfamily of peroxidase enzymes (4). generated in situ from glucose oxidase and glucose
While the molecular biology of M. tuberculosis resis-
was used. MtHPI-e was more effective at isoniazid-de-
tance to INH involving KatG-deficiency has been dem-
pendent radical production than EcHPI and HRP. Sim-
onstrated, the biochemical basis for resistance remains
ilar qualitative results were obtained by staining non-
less clear. INH is believed to affect several intracellular
denaturing polyacrylamide gels for activity with ni- troblue tetrazolium in the presence of isoniazid and
targets of mycolic acid synthesis (5), including an enoyl-
[acyl-carrier-protein] reductase (InhA) (6). Recently,
The absorbance spectrum of HRP exhibited changes during incubation with isoniazid and H2O2
Schultz and colleagues (7) have proposed a plausible
consistent with the formation of several typical reac-
mechanism of action for the drug based on in vitrotion intermediates, whereas the catalase – peroxidases
studies of purified KatG and InhA proteins which sug-
exhibited no distinct spectral changes. The results sug-
gest that INH must be oxidized in vivo to an electro-
gest that the sensitivity of M. tuberculosis to isoniazid
philic species by the catalase – peroxidase. The INH-
may be the result of isoniazid-dependent radical for-
derived electrophilic species then binds irreversibly to
mation by the catalase – peroxidase in the absence of
a cysteine residue near the active site of InhA, thereby
other catalase activities to remove substrate H2O2.
preventing assembly of mycolic acids leading to cyto-
᭧ 1995 Academic Press, Inc.
toxicity (7). If this scheme is correct, other as yet un-
Key Words: catalase – peroxidase; Mycobacterium tu-
identified cellular components may also be inactivated
berculosis; Escherichia coli; isoniazid; free radicals; ni-
or destroyed by KatG-generated free radicals through
troblue tetrazolium.
3 Abbreviations used: EcHPI, Escherichia coli catalase–peroxidase
hydroperoxidase I; MtHPI-e, Mycobacterium tuberculosis catalase –
1 This work was supported by an NSERC operating grant (OGP
peroxidase hydroperoxidase I – extended protein; HPII, catalase hy-
9600) to P.C.L. and an NSERC postgraduate research scholarship
droperoxidase II; HRP, horseradish peroxidase; InhA, Mycobacte-rium tuberculosis enoyl-[acyl-carrier-protein] reductase; INH, isoni-
2 To whom correspondence should be addressed. Fax: (204) 275-
cotinic acid hydrazide; SDS, sodium dodecyl sulfate; PAGE, poly-
7615. E-mail: [email protected].
acrylamide gel electrophoresis; NBT, nitroblue tetrazolium.
Copyright ᭧ 1995 by Academic Press, Inc.
All rights of reproduction in any form reserved.
COMPARATIVE OXIDATION OF ISONIAZID BY PEROXIDASES
INH oxidation. E. coli does not synthesize mycolic acid
Mechanism-based inactivation of HRP by radical spe-
and possesses a second monofunctional catalase en-
cies produced during incubation with phenylhydrazine,
zyme (HPII) which would remove H2O2 before it could
alkylhydrazines, and sodium azide has been shown
potentiate INH toxicity, possibly explaining why E. coli
previously (15). For catalase, catalase – peroxidase, and
is resistant to INH. Similarly, other INH-resistant My-
HRP, the reaction cycle incorporates reaction [1], but
cobacteria have an additional catalase, providing an
catalase enzymes preferentially reduce compound I via
explanation for their resistance. Initial attempts to
a one-step, two-electron reaction in which the substrate
characterize the role of INH in KatG-mediated cell tox-
donor is usually a second molecule of hydrogen perox-
icity were incomplete because crude cell extracts were
ide. In the case of HRP, at high H2O2 to donor substrate
used, but it was demonstrated that INH was taken up
concentration ratios, as well as in the absence of donor
by M. tuberculosis and oxidized (1).
substrates, the normal peroxidase cycle partitions at
Diaz and Wayne (8) and, subsequently, Gayathri-
compound II to react with H2O2 (16) or radical species
Devi and co-workers (9) reported the purification of
(17), yielding a reversibly inactivated HRP species
KatG from M. tuberculosis and demonstrated that
called compound III, or oxyperoxidase, at an oxidation
it had properties similar to horseradish peroxidase
state of VI. This species may further react with radical
(HRP), including optimum pH and activities in the
species or other donor molecules to regenerate com-
presence of various inhibitors and other reagents.
pound I (17). Additionally, compound I may react
The former report also showed that the catalase ac-
slowly under these conditions with H2O2 to yield either
tivity of KatG was inhibited in the presence of INH.
a second (irreversibly) inactivated form of the enzyme
More recently, it has been demonstrated that HRP
known as compound IV (16, 18), or to reduce compound
(10), extracts derived from M. tuberculosis (11), and
I back to resting (ferric) HRP in a catalase (two-elec-
human myeloperoxidase (12) can catalyze the INH-
tron) reaction (15), though the evidence for the latter
dependent generation of radical species. Purified,
plasmid-expressed M. tuberculosis KatG has also
In order to clarify the role of M. tuberculosis KatG
been used in mechanistic studies to confirm that oxi-
protein in INH toxicity, we have compared properties of
dized derivatives of INH generated by KatG include
a M. tuberculosis KatG protein with a carboxy-terminal
isonicotinic acid, isonicotinamide, and pyridine-4-
extension (MtHPI-e), EcHPI, and HRP. We have also
compared the ability of each enzyme to produce free
The general sequence of reactions for enzymatic
radicals and monitored the susceptibility of each en-
turnover of horseradish peroxidase with a hydrogen
zyme to inactivation, during incubation with INH in
donor in the presence of H2O2 is shown below:
the presence of low levels of H2O2. The results confirmthat the M. tuberculosis enzyme is more effective in
the generation of free radicals than either EcHPI or
HRP, explaining, in part, its role in INH-mediated tox-
MATERIALS AND METHODS
Common biochemicals, horseradish peroxidase (Types
II and IX), superoxide dismutase, and glucose oxidase (Type II) wereobtained from Sigma Chemical Co. Hydrogen peroxide (30%) andglucose were from Fisher Scientific, Ltd. LB media and molecular
where AH2 and rAH are reducing donor substrate and
biology reagents (enzymes and buffers) were from Gibco BRL. H2O2
free radical product, respectively. The numerals in pa-
concentration was determined immediately before applicable experi-
rentheses indicate formal oxidation states of HRP dur-
ing enzymatic turnover. The free radical products of
Expression and purification of enzymes.
the reaction may undergo various reactions depending
coli and M. tuberculosis were expressed from plasmids pBT22 (20)and pAH1. The latter was constructed by insertion of the 2.9-kb
on their chemistry, such as dimerization, reaction with
EcoRV – KpnI fragment of plasmid pYZ55 (2), containing the M. tu-
a neutral donor molecule, reaction with the enzyme
berculosis katG gene into pSK/ (Stratagene) where it was under
itself, or reduction of molecular oxygen to superoxide
control of the lac promoter. Restriction digestion, ligation, and clon-
(14, 15). Radical production under these conditions may
ing were as described by Maniatis et al. (21). Plasmids were trans-formed into E. coli UM262 [recA katG : : Tn10 pro leu rpsL hsdM hsdR
also be monitored by including a suitable electron ac-
endI lacY (22)], lacking both HPI and HPII. EcHPI was isolated as
ceptor such as nitroblue tetrazolium (NBT) in the reac-
described (23). MtHPI-e was isolated following basically the same
procedure with the following modifications: (1) catalase activity wasprecipitated at 35 and 40% of saturation with (NH4)2SO4, rather
4)2SO4 ; (2) the resuspended, pooled fractions from
(NH4)2SO4 precipitation were incubated for 45 min at 42ЊC and cen-
trifuged to remove additional precipitates prior to overnight dialysis;
and (3) pooled and dialyzed fractions recovered from the anion-ex-change column were loaded onto a Sephadex G-200 gel filtrationcolumn equilibrated with 50 mM potassium phosphate, pH 7.0, andeluted in 3-ml fractions. Catalase activity eluted with the first proteinpeak and fractions to be pooled were selected based on relative pro-tein purity estimated by electrophoresis on SDS – polyacrylamidegels. Pooled fractions were stored frozen (080ЊC) either in potassiumphosphate buffer or lyophilized. Lyophilized commercial HRP wasresuspended in 50 mM potassium phosphate buffer, pH 7.0, with nofurther purification and stored frozen (020ЊC).
Absorbance spectra, time courses, and perox-
idatic assays were performed using a Milton Roy MR3000 spectro-photometer. All experiments were performed at room temperaturein 1-ml quartz, semimicro cuvettes. Proteins were normally dilutedin 50 mM potassium phosphate buffer, pH 7.0, and the same bufferwas used as a reference. Activity assays and protein determination.
determined by the method of Rørth and Jensen (24), using a Gilson
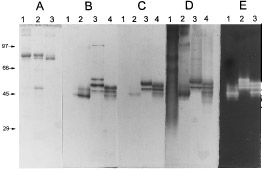
Polyacrylamide gel electrophoresis of HRP and the cata-
oxygraph equipped with a Clark electrode. One unit of catalase is
lase – peroxidases. Running conditions and procedures were as de-
defined as the amount that decomposes 1 mmol of H2O2 in 1 min at
scribed under Materials and Methods. MtHPI-e trypsin digests (30
37ЊC. Peroxidase activity was determined by the method described
min) were done at an enzyme/protease ratio of 100 (w/w) at room
in the Worthington Enzyme Catalogue (Worthington Chemical Co.),
temperature. Protein was stained with Coomassie brilliant blue, R-
modified to be carried out in 1-ml assay volumes. The average rate
250. (A) SDS-denaturing PAGE stained for protein. Estimated total
of H2O2 production from experiments employing the glucose/glucose
protein loaded was 25 mg for all lanes. High molecular weight mark-
oxidase system was estimated by determination of H2O2 concentra-
ers indicated by arrows. Lane 1, MtHPI-e; lane 2, MtHPI, trypsin
tion according to the method of Green and Hill (25) for duplicate
digest; lane 3, EcHPI. (B – D) Nondenaturing PAGE stained for pro-
or triplicate samples, at 5-min intervals, allowing 5 min for color
tein (B), peroxidatic oxidation of INH with NBT (C), or peroxidatic
development before making individual absorbance readings at 505
oxidation of diaminobenzidine (D). Lane 1, HRP (type IX); lane 2,
nm. Protein was estimated by the methods described by Layne (26).
EcHPI; lane 3, MtHPI; lane 4, MtHPI-e, trypsin digest. (E) Nondena-
cm01 for the monoformazan product from the
turing PAGE stained for catalase. Lane 1, EcHPI; lane 2, MtHPI-e;
Polyacrylamide gel electrophoresis and visualization of enzymatic
Gel electrophoresis of purified proteins was carried out
under denaturing conditions on SDS – polyacrylamide gels as pre-viously described (28, 29). Gel electrophoresis was carried out under
to be larger due to a polypeptide extension. The N-
nondenaturing conditions according to Davis (30), except in pH 8.1
terminal sequence of the purified protein for the first
Tris – HCl. For all electrophoretic procedures, 15 1 15-cm slab gelswere run in a Protean II electrophoretic chamber (Bio-Rad). Follow-
9 amino acids corresponded to that predicted from the
ing electrophoresis, peroxidase activity was visualized by the method
DNA sequence of M. tuberculosis katG, indicating that
of Gregory and Fridovich (31) and catalase was visualized as de-
the polypeptide was larger due to a C-terminal exten-
scribed by Clare et al. (32), but using 20 mM H2O2 for better contrast.
sion, rather than an N-terminal LacZ fusion. The rea-
Peroxidase-mediated oxidation of INH was visualized by soaking thegels in 200 ml 50 mM potassium phosphate, pH 7.0, containing 274
son for the longer than expected terminal end is cur-
mg INH, 50 mg NBT, and 60 ml 30% H2O2. Color development was
rently under investigation. Two possibilities include
usually complete between 30 min and 1 h, after which gels were
differences from the published sequence or aberrant
rinsed with distilled water and soaked in 7% acetic acid, 1% glycerol
translation of the G-C rich mRNA in E. coli. Transla-
overnight before mounting. Bands usually intensified slightly during
tion ending at the next termination site would give rise
incubation. Incubation of gels for 16 h in the staining solution fromwhich NBT was omitted also resulted in band development, but the
to a 49-amino-acid extension at the carboxyl-terminal
bands were much weaker than those seen using the complete stain-
of MtHPI. Under nondenaturing conditions (Fig. 1B),
EcHPI exhibited the normal double band of charge iso-forms corresponding in size to a dimer, while MtHPI-
RESULTS AND DISCUSSION
e exhibited three bands also presumed to be chargeisoforms. Fortuitously, the predicted sequence of
Characteristics of MtHPI-e and EcHPI Compared
MtHPI-e contains several closely spaced arginine and
lysine residues in the region immediately following the
Purified EcHPI and MtHPI-e were analyzed by elec-
usual final residue of the MtHPI which proved to be a
trophoresis on polyacrylamide gels (Fig. 1). Under de-
target for trypsin cleavage to generate a nearly native
naturing conditions (Fig. 1A), both proteins exhibited
MtHPI. As shown in Figs. 1A and 1B, a protein band
a predominant single band of protein with apparent
with mobility comparable to EcHPI under both dena-
molecular masses of 82 kDa for EcHPI and 89 kDa for
turing and nondenaturing conditions was obtained
MtHPI-e. As unmodified EcHPI and MtHPI polypep-
from cleavage of MtHPI-e with trypsin.
tides have over 50% sequence identity and differ by
Staining for peroxidase activity in polyacrylamide
only 11 amino acids in length, MtHPI-e was inferred
gels may be accomplished by several techniques (31,
COMPARATIVE OXIDATION OF ISONIAZID BY PEROXIDASES
an inactive component, its presence should not affectthe peroxidase/catalase ratio or any of the subsequentresults. It should be pointed out that previous purifica-tions of wild-type MtHPI have reported A408/280 ratiosof 1.0 (9) and 0.37 (8); the latter value being the sameas that for our purified MtHPI-e. Furthermore, the util-ity of a fusion protein has been demonstrated by Zhanget al. (2), who have previously shown a LacZ – MtHPIfusion to confer INH susceptibility on E. coli. This fu-sion protein has recently been reported purified, havinga specific catalase activity of 494 U/mg and exhibitingthree isoforms in nondenaturing PAGE (34). Rate and Extent of INH-Dependent Generation of
Absorbance spectra of HRP and the catalase – peroxidases. Free Radicals by HRP are Dependent on H2O2
Absorbance spectra of MtHPI-e (solid line), EcHPI (dotted line), and
HRP (dashed line) hemoproteins in the Soret and visible (inset) re-gions. Samples were suspended in potassium phosphate buffer, pH
HRP and myeloperoxidase catalyze a reaction depen-
7.0. Spectra obtained were factor adjusted to absorbance equality at
dent on the presence of INH and H2O2 that may be
the Soret maxima for comparison purposes. The scale was expandedin the inset by the factor shown.
followed by reduction of NBT to its purple formazanproduct (10, 11). Shoeb and colleagues (10) also ob-served that the presence of exogenous catalase caused
33). An alternative peroxidase stain was suggested by
an increase in the rate of NBT reduction by HRP and
the observation of Shoeb et al. (10) that HRP reacted
a faster termination of the reaction rather than the
with INH and H2O2 to produce radical products that
expected cessation of reaction as catalase removed the
could reduce NBT to its purple formazan product. NBT
substrate H2O2. They also confirmed that the presence
has been used for other activity stains on polyacryl-
of superoxide dismutase did not inhibit NBT reduction
amide gels including the superoxide dismutase stain
in the INH-dependent reaction catalyzed by HRP.
(31). Purified EcHPI, MtHPI-e, and HRP were stained
To determine if the catalase effect on HRP was solely
with NBT – INH (Fig. 1C), revealing bands that corres-
the result of the lower [H2O2], we varied the initial
ponded in location and intensity to the peroxidase
[H2O2] and followed the INH/H2O2-dependent NBT re-
bands visualized using the diaminobenzidene stain
duction by HRP (Fig. 3A). Two distinct patterns of NBT
(Fig. 1D). As expected the catalase activity (Fig. 1E) in
reduction were observed depending on the [H2O2]. At
the two HPI proteins comigrated with the peroxidase
[H2O2] up to 100 mM, there was a rapid reaction rate
which ceased after a time proportional to the initial
The absorbance spectra of EcHPI, MtHPI-e, and
[H2O2]. At higher [H2O2], the initial fast phase became
HRP in the Soret and visible regions of the spectrum
progressively shorter and, eventually, indistinguish-
are shown in Fig. 2. Very similar spectra were obtained
able from the second, slower phase. This change in rate
for EcHPI and MtHPI-e with clear maxima at 408 and
at higher [H2O2] is presumably a result of a progressive
500 nm although the Soret peak of MtHPI-e was
inactivation of the enzyme as [H2O2] increases in the
slightly broader and the 639 nm band of EcHPI (and
presence of donor substrate (15). The maximal initial
HRP) was positioned at 628 nm in MtHPI-e. By con-
reaction rate of NBT reduction with 100 mM H2O2 added
trast, the Soret band of HRP was located at 404 nm,
was estimated to be 9 nmol/min/ml. [NBT] was varied
and there were more sharply defined maxima at 510
from 20 to 600 mM to determine whether efficiency of
the reaction could be improved by reducing the proba-
The specific activities for the various enzymes listed
bility of any side reactions of the radicals generated.
in Table I reveal that the peroxidase/catalase ratio for
Apart from a marginal increase in the rate of the initial
MtHPI-e is 50% higher than the similar ratio for
phase of reaction at lower [NBT] (20 mM), varying the
EcHPI. The possible importance of this in H2O2-depen-
[NBT] had no effect over the concentration range
dent radical production will be discussed below. The
tested. In addition, the reaction was monitored in the
A408/280 ratios are also presented in Table I and reveal
presence of superoxide dismutase (1.5 U), added both
a lower ratio for MtHPI-e than for EcHPI, suggesting
prior to and following the initiation of NBT reduction,
that approximately 25% of the MtHPI-e lacks heme.
to ensure that superoxide anion radical was not a sig-
This is consistent with the minor upper protein band
nificant proportion of the radicals produced. No decline
in the nondenaturing gels (Fig. 1B) that is inactive in
in reaction rate was observed in the presence of super-
both the catalase- and the peroxidase-stained gels. As
oxide dismutase for these experimental controls.
Selected Properties of the Catalase – Peroxidases and Comparison with HRP
a Ratio reported for HRP is A404/280.
The effect of INH concentration on NBT reduction at
MtHPI-e, EcHPI, and HRP Have Different Rates of
an initial [H2O2] of 250 mM is shown in Fig. 3B. The
NBT Reduction under Conditions of Constant H2O2
initial rate of reduction was proportional to INH con-
centration and there was no evidence of termination
Our initial attempts at evaluating the capacity of
of the reduction at higher INH concentrations. This
MtHPI-e and EcHPI to mediate the oxidation of INH,
suggests that the termination of radical production
via the NBT reduction assay used for HRP, were con-
seen for HRP observed in Fig. 3B is dependent upon
founded by the catalase activities of the MtHPI-e and
[H2O2], not the ratio of [H2O2] to [donor].
the EcHPI obscuring their peroxidatic activities underconditions of single peroxide additions where the cata-lase activity would rapidly remove H2O2 from the me-dium. In order to generate a constant source of H2O2,glucose oxidase and glucose were added to the system,resulting in the generation of H2O2 at a rate of approx-imately 7 nmol/ml/min. The results of exposingMtHPI-e, EcHPI, and HRP to this constant peroxideflux in the presence of INH and NBT are shown inFig. 4. It is apparent that MtHPI-e was much moreeffective than EcHPI and HRP in mediating INH oxi-dation and NBT reduction compared to catalyzing per-oxidation of o-dianisidine. Comparison of the kineticsfor INH oxidation of the catalase – peroxidases basedon the amount of NBT reduced under these assay con-ditions yielded Kms of 1.24 and 1.49 mM and kcat values
Time courses for HRP-mediated oxidation of INH followed
by NBT reduction. Experiments done at room temperature in semi-micro quartz cuvettes containing 1-ml final volumes of NBT (0.2 mM),HRP (30 mg), and potassium 50 mM phosphate buffer, pH 7.0. (A) In
Comparison of INH oxidation rates of catalase – peroxidases
the presence of 9 mM INH, H2O2 addition initiated the reaction at the
and HRP. Experiments done at room temperature in semimicro
time indicated by the arrow at the following initial concentrations: (a)
quartz cuvettes containing 1-ml final volumes of NBT (0.2 mM), INH
5 mM, (b) 10 mM, (c) 50 mM, (d) 100 mM, (e) 1 mM, (f) 500 mM, (g) 250
(9 mM), glucose oxidase (5 mg), and either 1 U peroxidase (0.2 mg)
mM. (B) H2O2 addition (250 mM) initiated the reaction (arrow) in the
of EcHPI (dotted line), 1 U peroxidase (0.2 mg) of HRP (broken line),
presence of the following initial concentrations of INH: (a) 87.5 mM,
or 1 U peroxidase (0.2 mg) of MtHPI-e (solid line). Addition of glucose
(b) 440 mM, (c) 875 mM, (d) 4.4 mM, (e) 17.5 mM, and (f) 52.5 mM.
(4 mM) initiated the reaction at the time indicated by the arrow.
COMPARATIVE OXIDATION OF ISONIAZID BY PEROXIDASES
of 0.092 and 0.58 s01 for EcHPI and MtHPI, respec-
properties, which we believe provide good support for
tively. The rate of NBT reduction by MtHPI-e in-
the likelihood that they produce similar radical species.
creased with the addition of more enzyme, indicating
Two further explanations for the greater radical pro-
that the rate of H2O2 formation was not limiting under
duction by MtHPI-e include the possibility that INH is
these conditions. However, the addition of more than
a more effective substrate for MtHPI than for EcHPI,
100 peroxidase units of HRP did not elicit further in-
and the possibility that EcHPI is more susceptible to
creases in the initial rate of reaction. Similar to the
inactivation by INH-derived radicals. The latter possi-
slowing of the NBT reduction rate upon single addi-
tion of H2O2 (Fig. 3), the rates of NBT reduction alsodeclined over time in the presence of a continuous
Spectral Evaluation of INH Oxidation Mediated by
flux of H2O2. As the rate of generation of H2O2 was
confirmed to be stable for at least 15 min under theconditions employed, this phenomenon may be attrib-
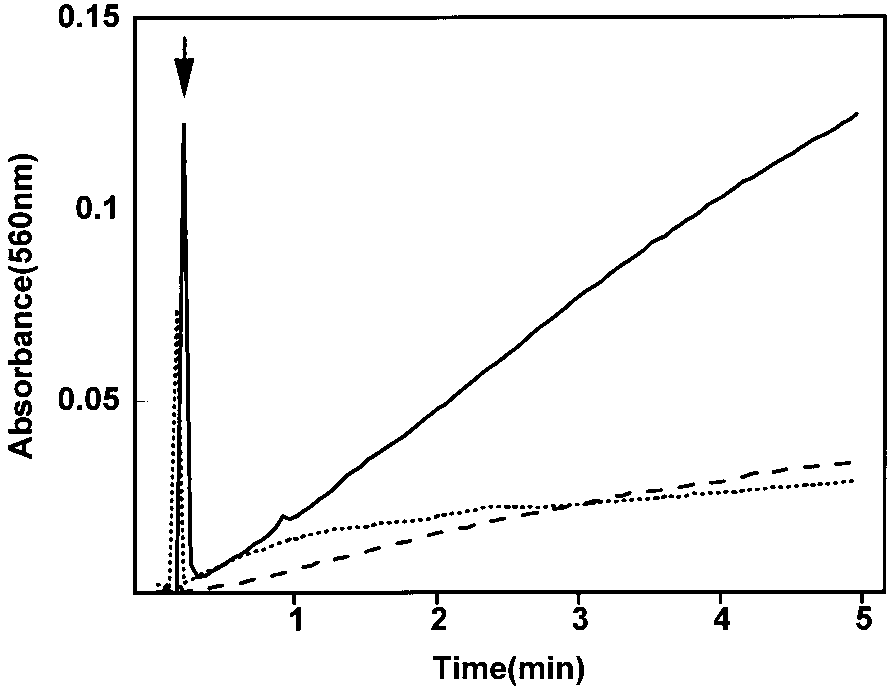
The incubation of the compound II form of HRP
uted to either a progressive inhibition of the radical
with INH resulted in spectral shifts, including a
generating reaction(s) or the establishment of a steady
bleaching of the Soret band, suggestive of heme split-
state of formation for the radical species present.
ting which would be synonymous with inactivation
The finding that all three enzymes mediate the oxi-
(10). It was unclear whether the decline in radical
dation of INH to yield radical species is not surprising
production in the presence of a constant flux of H2O2
in view of their common peroxidase activity. What is
(Fig. 4) was the result of the establishment of a
of more immediate interest is the finding that the
steady state or of a progressive inhibition of the reac-
MtHPI-e supports a higher rate of radical generation
tion(s) involved in the generation of radicals possibly
than do either HRP or EcHPI under conditions of con-
by enzyme inactivation. To address this, we moni-
stant peroxide flux. The catalytic activity responsible
tored both the spectra and the peroxidase activities
for the INH-dependent generation of radicals captured
of MtHPI-e, EcHPI, and HRP before and after expo-
by NBT is not directly proportional to the o-dianisidine
sure of the proteins to the peroxide-generating sys-
peroxidase activity among the three enzymes. MtHPI
tem, in the presence and absence of INH.
has a 10% higher peroxidase specific activity than
The absorption spectra of the enzymes before and
EcHPI, but causes NBT reduction to a threefold higher
after a 10-min incubation with the constant flux of
level, which could, in part, explain the ability of MtHPI
H2O2 from glucose oxidase/glucose with and without
to enhance INH sensitivity of Mycobacteria and confer
INH are shown in Fig. 5. In the absence of INH, there
INH sensitivity on E. coli (2). Early in the reaction
is a red shift of the Soret band of HRP to 418 nm,
there will be a partition of the enzyme between cata-
combined with the disappearance of the band at 639
latic and peroxidatic modes at compound I (the en-
nm and appearance of maxima at 550 and 580 nm (Fig.
zyme – H2O2 complex) resulting in the eventual estab-
5A). These spectral shifts are indicative of the forma-
lishment of a steady state. In the case of MtHPI-e, the
tion of compound III or oxyperoxidase, a reversibly in-
proportion of peroxidatic (and derived radical forma-
activated form of the enzyme that is typically observed
tion) to catalatic activity will be greater than for
at high peroxide – enzyme ratios or in the presence of
EcHPI. However, a simple disproportion of peroxidatic
certain electron donors (16). In the presence of INH,
to catalatic activities is not a sufficient explanation be-
however, the Soret band was markedly reduced in in-
cause an E. coli HPI mutant with a peroxidatic to cata-
tensity as well as red-shifted to 418 nm, and a broad
latic activity ratio similar to MtHPI-e (Table I, 22) was
band at 530 nm with a shoulder at 550 nm, as well as
no more effective in the INH-dependent generation of
a peak at 670 nm, appeared. The maximum at 670 nm
radicals than the wild-type MtHPI. It may also be ar-
is indicative of the formation of compound IV or P670,
gued that the three enzymes examined mediate produc-
a second, irreversibly inactivated form of the enzyme
tion of different types of radical species, resulting in
(15), while the peak with the shoulder at 530 nm sug-
different rates of NBT reduction. The products of reac-
gests the presence of HRP compound II, or ferryl form
tion of INH with MtHPI have been identified as isoni-
of the enzyme. The decline in the intensity of the Soret
cotinic acid, isonicotinamide, and pyridine-4-carboxal-
peak is indicative of a major perturbation of the elec-
dehyde, consistent with a scheme of enzymatic oxida-
tronic structure of the heme, attributable to either
tion of INH potentially involving hydrazide radical,
heme modification or degradation. Indeed, the heme of
diazenyl radical, and an acyl radical (13). As NBT re-
HRP has been shown to be susceptible to reaction with
duction cannot be used to distinguish between these
phenylhydrazine-derived radicals, resulting in a simi-
radicals, the relative reaction rates of the individual
radicals with the acceptor cannot be estimated. How-
Because a fast peroxidatic mode of reaction was not
ever, the catalase – peroxidases examined in this study
observed for HRP and because compound III and IV
exhibit many similarities in their physicochemical
forms of the enzyme were observed, the involvement of
INH as an electron donor may therefore result in thepartition of HRP compound II into the oxidatic modeand to accelerate the decay of compound III to com-pound I for reinitiation of the cycle. This may protectthe HRP from further inactivation but may also be thereason for the significantly slower generation of radi-cals from INH compared to the MtHPI-e protein.
The spectra obtained for EcHPI and MtHPI-e (Figs.
5B and 5C) in presence of the peroxide-generating sys-tem, with and without INH, did not show spectralchanges as distinct as those seen for HRP. In the ab-sence of INH, the Soret peak and the band at 639 nmintensified slightly, while the 500-nm broad band de-clined in intensity. A similar type of spectral shift hasbeen observed in the visible region of the spectrumfor purified recombinant ascorbate peroxidase, when astoichiometric amount of H2O2 was added to the en-zyme (36). The presence of INH resulted in the forma-tion of a broad band around 510 nm and a slight declinein the intensity of the peak at 639 nm for both enzymes. MtHPI-e showed no change in intensity of the Soretpeak, whereas EcHPI showed a slight decline in theintensity of the same peak. These spectral changes sug-gest that the hemes of the catalase – peroxidases do notundergo major electronic perturbations or modificationduring reaction with H2O2 and INH under the condi-tions chosen. It should be pointed out, however, thatthe partition between the one-step, two-electron reduc-tion of compound I via the catalatic cycle and the two-step, one-electron reductions of compound I via the per-oxidatic cycle favors the former cycle’s steady-stateintermediate(s). This caveat requires these spectral re-sults to be interpreted with appropriate caution.
As noted above, one possible explanation for the
lower rate of INH-dependent radical production by
Peroxidatic Activities of the Catalase – Peroxidases
and HRP during Incubation in the Presence
HRP spectral changes during incubation with INH in pres-
ence of constantly generated H2O2. Conditions as described in thelegend to Fig. 4. Spectra for each hemoprotein were obtained prior
to initiation of H2O2 generation (solid lines), 10 min following addi-
tion of glucose (dotted lines), and 10 min following addition of glucose
in presence of INH (9 mM) (broken lines). Insets show visible regions
of the spectrum at scales expanded by the factors shown. (A) Spectra
of HRP (30 mg). (B) Spectra of MtHPI-e (0.2 mg). (C) Spectra of EcHPI
Note. Incubations were done under conditions described in the
an oxidatic mode may be inferred (17). The oxyperoxi-
legend to Fig. 5. At the times indicated, aliquots were withdrawn
dase intermediate of the oxidase cycle has an extremely
from the incubation mixtures and assayed for peroxidatic activity as
slow rate of decay and is usually formed at high [H2O2],
outlined under Materials and Methods.
and its function has been hypothesized to rescue the
a Data are expressed as the percentage of o-dianisidine activity
enzyme from irreversible inactivation. The presence of
remaining compared to the activity before the addition of glucose.
COMPARATIVE OXIDATION OF ISONIAZID BY PEROXIDASES
TABLE III
role of a catalase – peroxidase such as MtHPI, which is
Catalase Activities of the Catalase – Peroxidases during
capable of producing high levels of radical species via
Incubation in the Presence of Constantly Generated H2O2
oxidation of INH, would therefore be as an efficientprodrug activator in the envisioned scheme. However,
even in the absence of a drug target, it is conceivablethat higher intracellular levels of INH-derived radical
species alone would also have deleterious effects.
The biochemical bases for INH resistance in a sig-
nificant proportion of M. tuberculosis clinical isolates
Note. Incubations were done under conditions described in the
legend to Fig. 5. At the times indicated, aliquots were withdrawn
is thus also apparently realized. It is estimated that
from the incubation mixtures and assayed for catalase activity as
25 – 50% of INH-resistant M. tuberculosis isolates are
outlined under Materials and Methods. Activities assayed in absence
also catalase – peroxidase negative, and that another
20 – 25% of resistant isolates have mutations in the
Data are expressed as the percentage of catalase activity re-
maining compared to the activity before the addition of INH and
inhA locus (6). It would therefore appear that either
modification of the gene for the target protein such thatInhA fails to covalently bind to INH-derived radicals,or inactivation of the gene for the prodrug activator
EcHPI is that the enzyme is more sensitive to inactiva-
(MtHPI) such that production of INH-derived radicals
tion by INH-derived radicals. While the lack of signifi-
is eliminated or reduced, are two of the major determi-
cant spectral changes argued against this possibility,
nants in strains of M. tuberculosis acquiring INH-resis-
changes in the o-dianisidine activity were assayed dur-
ing incubations with a constant flux of H2O2 in presenceand absence of INH (Table II). There was a slow INH-
ACKNOWLEDGMENTS
dependent drop in HRP peroxidase activity of 24% over
We thank Stewart Cole and Judah Rosner for the gifts of plasmid
the reaction time of 10 min. INH alone caused a sig-
nificant inhibition of both MtHPI-e and EcHPI but thetwo enzymes differed in that the activity of MtHPI-e
REFERENCES
recovered slowly during incubation with H2O2. Perox-
1. Winder, F. G. (1982) in The Biology of the Mycobacteria
ide alone had no effect on MtHPI-e but caused a slow
(Ratledge, C., and Stanford, J., Eds.), Vol. 1, pp. 353 – 438, Aca-
inactivation of EcHPI. Similarly, when catalase activ-
ity of the HPI enzymes was assayed during incubation
2. Zhang, Y., Heym, B., Allen, B., Young, D., and Cole, S. (1992)
under these conditions (Table III), INH caused greater
Nature (London) 358, 591 – 593.
inhibition of EcHPI than MtHPI-e over the 10-min in-
3. Altamirano, M., Marostenmaki, J., Wong, A., FitzGerald, M.,
cubation period, while the activity of both enzymes re-
Black, W. A., and Smith, J. A. (1994) J. Infect. Dis. 169, 1162 –
mained unaffected in presence of continuously gener-
4. Welinder, K. G. (1992) Curr. Opinion Struct. Biol. 2, 388 – 393.
2O2 alone. This result agrees with a previous
study of MtHPI which demonstrated that INH inhib-
5. Banerjee, A., Dubnau, E., Quemard, A., Balasubramanian, V.,
Sun Um, K., Wilson, T., Collins, D., de Lisle, G., and Jacobs,
ited catalase activity of the enzyme (7). Thus, the
W. R. (1993) Science 263, 227 – 230.
MtHPI-e enzyme does seem to be more resistant to INH
6. Dessen, A., Quemard, A., Blanchard, J. S., Jacobs, W. R., and
Sacchettini, J. C. (1995) Science 267, 1638 – 1641.
We are left with the conclusion that MtHPI-e is
7. Johnsson, K., King, D. S., and Schultz, P. G. (1995) J. Am. Chem.
intrinsically more effective at INH-dependent genera-
Soc. 117, 5009 – 5010.
tion of radicals than EcHPI. This may in part be ex-
8. Diaz, G. A., and Wayne, L. G. (1974) Am. Rev. Respir. Dis. 110,
plained by INH being a better substrate for MtHPI
than for the EcHPI and by the MtHPI being less sensi-
9. Gayathri-Devi, B., Shaila, M. S., Ramakrishnan, and Gopina-
tive to INH-mediated inactivation than the EcHPI. The
than, K. P. (1976) Biochem. J. 149, 187 – 197.
significance of our results is enhanced by the recently
10. Shoeb, H. A., Bowman, B. U., Ottolenghi, A. C., and Merola,
A. J. (1985) Antimicrob. Agents Chemother. 27, 399 – 403.
proposed mechanism of action of INH against M. tuber-
11. Shoeb, H. A., Bowman, B. U., Ottolenghi, A. C., and Merola,
culosis (7), in which INH is a prodrug that is activated
A. J. (1985) Antimicrob. Agents Chemother. 27, 404 – 407. in vivo by MtHPI to an electrophilic species. The elec-
12. van der Walt, B. J., van Zyl, J. M., and Kriegler, A. (1994) Int.
trophilic species subsequently reacts with the Cys-243
J. Biochem. 26, 1081 – 1093.
residue of an enoyl-[acyl-carrier-protein] reductase (6),
13. Johnsson, K., and Schultz, P. G. (1994) J. Am. Chem. Soc. 116,
the putative drug target, resulting in its irreversible
inactivation, thereby impairing an intermediate step
14. Dunford, H. B. (1991) in Peroxidases in Chemistry and Biology
in fatty acid synthesis. This ultimately results in the
(Everse, J., Everse, K. E., and Grisham, M. B., Eds.), Vol. II, pp.
failure of mycolic acid synthesis and cytotoxicity. The
1 – 24, CRC Press, Boca Raton, FL.
15. Baynton, K. J., Bewtra, J. K., Biswas, N., and Taylor, K. E.
25. Green, M. J., and Hill, H. A. O. (1984) Methods Enzymol. 105,
(1994) Biochim. Biophys. Acta 1206, 272 – 278.
26. Layne, E. (1957) Methods Enzymol. 3, 447 – 454.
16. Nakajima, R., and Yamazaki, I. (1987) J. Biol. Chem. 262, 2576 –
27. Auclair, C., and Voisin, E. (1985) in CRC Handbook of Methods
for Oxygen Radical Research (Greenwald, R. A., Ed.), pp. 123 –
17. Saikumar, P., Swaroop, A., Ramakrishna Kurup, C. K., and Ra-
masarma, T. (1994) Biochim. Biophys. Acta 1204, 117 – 123.
28. Laemmli, U. K. (1970) Nature (London) 227, 680 – 685.
18. Arnao, M. B., Acosta, M., del Rı´o, J. A., Varo´n, R., and Garcı´a-
29. Weber, K., Pringle, J. R., and Osborn, M. (1972) Methods Enzy-
Ca´novas, F. (1990) Biochim. Biophys. Acta 1041, 43 – 47. mol. 26, 3 – 27.
19. Nelson, D. P., and Kiesow, L. A. (1972) Anal. Biochem. 49, 474 –
30. Davis, B. J. (1964) Ann. N. Y. Acad. Sci. 121, 404 – 427.
31. Gregory, E. M., and Fridovich, I. (1974) Anal. Biochem. 58, 57 –
20. Triggs-Raine, B. L., and Loewen, P. C. (1987) Gene 52, 121 – 128.
32. Clare, D. A., Duong, M. N., Darr, D., and Archibald, F. (1984)
21. Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular
Anal. Biochem. 140, 532 – 537.
Cloning. A Laboratory Manual, Cold Spring Harbor Press, Cold
33. Wayne, L. G., and Diaz, G. A. (1986) Anal. Biochem. 157, 89 –
22. Loewen, P. C., Switala, J., Smolenski, M., and Triggs-Raine,
34. Nagy, J. M., Cass, A. E. G., and Brown, K. A. (1995) Biochem.
B. L. (1990) Biochem. Cell Biol. 68, 1037 – 1044. Soc. Trans. 23, 152S.
23. Loewen, P. C., and Switala, J. (1986) Biochem. Cell Biol. 64,
35. Ator, M. A., and Ortiz de Montellano, P. R. (1987) J. Biol. Chem.262, 1542 – 1551.
24. Rørth, M., and Jensen, P. K. (1967) Biochim. Biophys. Acta 139,
36. Patterson, W. R., and Poulos, T. L. (1994) J. Biol. Chem. 269,
Mittel gegen Seekrankheit („Seasickness remedies“, von Professor Noel Dilly, aus: ‚Adlard Coles’ Heavy Weather Sailing, 6th edition 2008 – Übersetzung aus dem Englischen durch Dr. med. Jens Kohfahl) Hoffentlich hat man sich so umfassend und rechtzeitig auf schweres Wetter vorbereitet, dass es keiner großen Anstrengung mehr bedarf, wenn es wirklich zu wehen anfängt. Man kann je
Fisher, Jill A. & Lorna M. Ronald (2008) “Direct-to-Consumer Responsibility: Medical Neoliberalism in Pharmaceutical Advertising and Drug Development.” In S.M. Chambré & M. Goldner (Eds.), Patients, Consumers and Civil Society (Advances in Medical Sociology, Volume 10) (pp. 29-51). Brighton, UK: Emerald Publishing. Definitive version can be accessed here: http://www.emeraldinsight.com
 and (3) pooled and dialyzed fractions recovered from the anion-ex-change column were loaded onto a Sephadex G-200 gel filtrationcolumn equilibrated with 50 mM potassium phosphate, pH 7.0, andeluted in 3-ml fractions. Catalase activity eluted with the first proteinpeak and fractions to be pooled were selected based on relative pro-tein purity estimated by electrophoresis on SDS – polyacrylamidegels. Pooled fractions were stored frozen (080ЊC) either in potassiumphosphate buffer or lyophilized. Lyophilized commercial HRP wasresuspended in 50 mM potassium phosphate buffer, pH 7.0, with nofurther purification and stored frozen (020ЊC).
and (3) pooled and dialyzed fractions recovered from the anion-ex-change column were loaded onto a Sephadex G-200 gel filtrationcolumn equilibrated with 50 mM potassium phosphate, pH 7.0, andeluted in 3-ml fractions. Catalase activity eluted with the first proteinpeak and fractions to be pooled were selected based on relative pro-tein purity estimated by electrophoresis on SDS – polyacrylamidegels. Pooled fractions were stored frozen (080ЊC) either in potassiumphosphate buffer or lyophilized. Lyophilized commercial HRP wasresuspended in 50 mM potassium phosphate buffer, pH 7.0, with nofurther purification and stored frozen (020ЊC).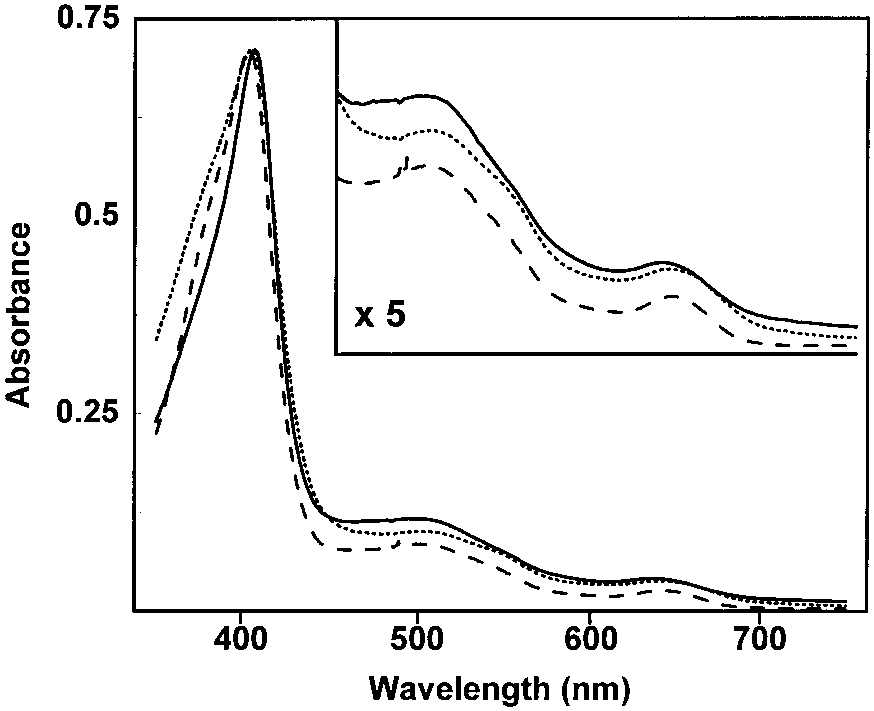 COMPARATIVE OXIDATION OF ISONIAZID BY PEROXIDASES
an inactive component, its presence should not affectthe peroxidase/catalase ratio or any of the subsequentresults. It should be pointed out that previous purifica-tions of wild-type MtHPI have reported A408/280 ratiosof 1.0 (9) and 0.37 (8); the latter value being the sameas that for our purified MtHPI-e. Furthermore, the util-ity of a fusion protein has been demonstrated by Zhanget al. (2), who have previously shown a LacZ – MtHPIfusion to confer INH susceptibility on E. coli. This fu-sion protein has recently been reported purified, havinga specific catalase activity of 494 U/mg and exhibitingthree isoforms in nondenaturing PAGE (34).
COMPARATIVE OXIDATION OF ISONIAZID BY PEROXIDASES
an inactive component, its presence should not affectthe peroxidase/catalase ratio or any of the subsequentresults. It should be pointed out that previous purifica-tions of wild-type MtHPI have reported A408/280 ratiosof 1.0 (9) and 0.37 (8); the latter value being the sameas that for our purified MtHPI-e. Furthermore, the util-ity of a fusion protein has been demonstrated by Zhanget al. (2), who have previously shown a LacZ – MtHPIfusion to confer INH susceptibility on E. coli. This fu-sion protein has recently been reported purified, havinga specific catalase activity of 494 U/mg and exhibitingthree isoforms in nondenaturing PAGE (34).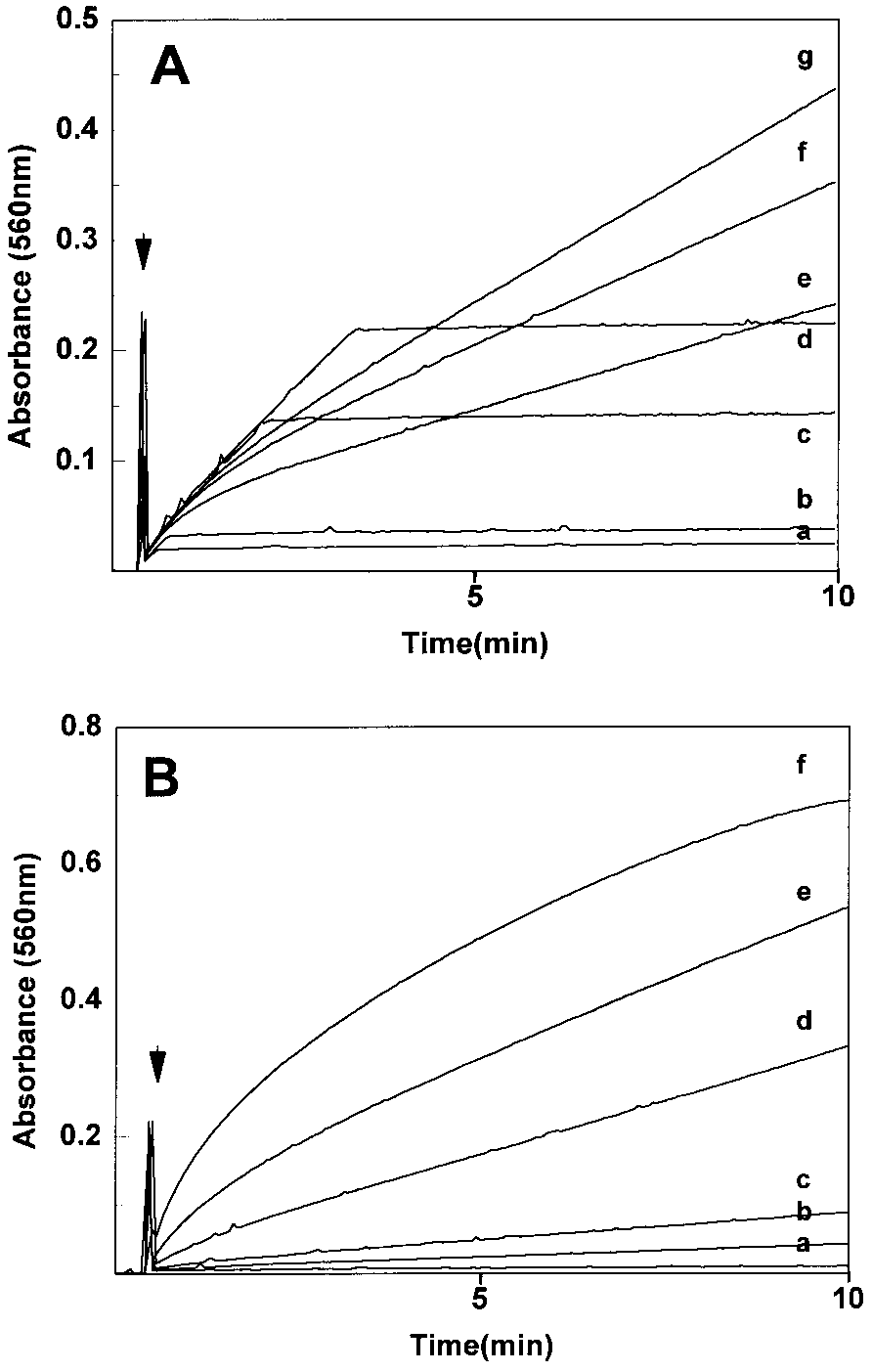
 Selected Properties of the Catalase – Peroxidases and Comparison with HRP
a Ratio reported for HRP is A404/280.
Selected Properties of the Catalase – Peroxidases and Comparison with HRP
a Ratio reported for HRP is A404/280.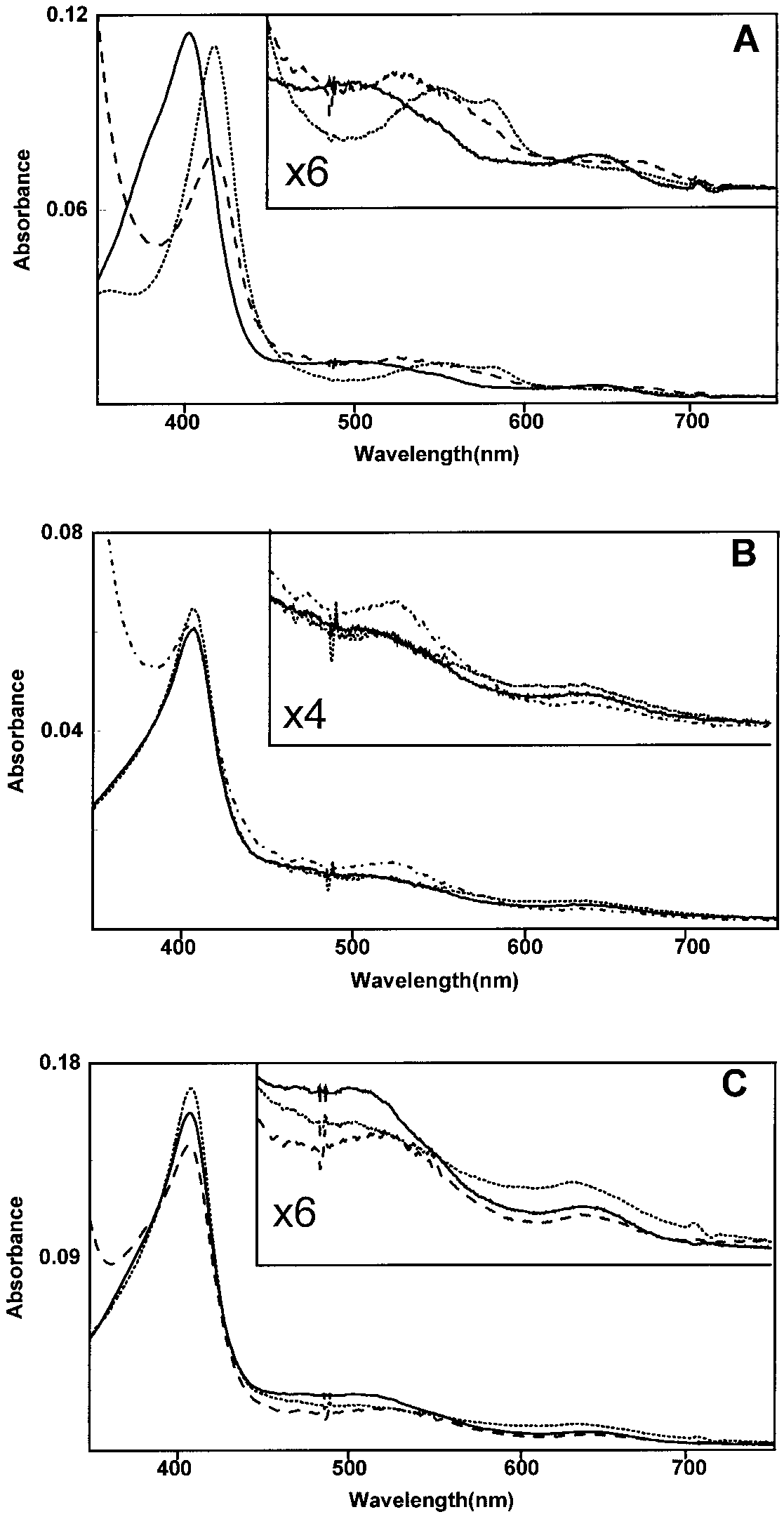 INH as an electron donor may therefore result in thepartition of HRP compound II into the oxidatic modeand to accelerate the decay of compound III to com-pound I for reinitiation of the cycle. This may protectthe HRP from further inactivation but may also be thereason for the significantly slower generation of radi-cals from INH compared to the MtHPI-e protein.
INH as an electron donor may therefore result in thepartition of HRP compound II into the oxidatic modeand to accelerate the decay of compound III to com-pound I for reinitiation of the cycle. This may protectthe HRP from further inactivation but may also be thereason for the significantly slower generation of radi-cals from INH compared to the MtHPI-e protein.