Practical medical trials have confirmed the maximum effect of Levitra, whith a positive effect being observed in 99 of 100 patients priligy uk Our internet store sells medicines produced in Indian pharmaceutical factories under licensed agreement.
Doi:10.1016/j.aquaculture.2003.09.003
Disease resistance of Pacific white shrimp,
Litopenaeus vannamei, following the dietary
administration of a yeast culture food supplement
Joseph E. Burgents, Karen G. Burnett*, Louis E. Burnett
Grice Marine Laboratory, College of Charleston, 205 Fort Johnson Road, Charleston, SC 29412, USA
Received 19 February 2003; received in revised form 20 August 2003; accepted 2 September 2003
A yeast culture feed supplement (Diamond V XP Yeast CultureR, Diamond V Mills, Cedar
Rapids, Iowa [IA]) was assessed for its impact on disease resistance in the Pacific white shrimp,Litopenaeus vannamei. Animals were fed a standard shrimp pellet diet supplemented with 0%(control with 1% grain carrier), 0.5% (with 0.5% carrier), or 1.0% XP daily for 4 weeks. To assessresistance to bacterial disease, at 1-week intervals 21 shrimp (0.5 – 2.5 g) from each test diet wereinjected intramuscularly with an LD50 dose (2.0 Â 105/g body weight) of a gram-negative shrimppathogen, Vibrio sp. 90-69B3. Survival was monitored every 4 h for 48-h post injection. Each week,three independent bacterial challenges were performed for each diet and the results expressed as themean percent survival F standard error (S.E.). A two-way analysis of variance (ANOVA) showed asignificant effect of diet ( p = 0.003, df = 31), but not duration of feeding, on survival. A one-wayANOVA showed no differences among the treatment groups after 1 or 2 weeks. After 3 weeks, themean survival of 1% XP-fed shrimp (74.2 F 1.4%) was significantly higher than that of controls(42.9 F 5.5%), while mean survival of shrimp fed 0.5% XP (54.8 F 11.9%) was not significantlydifferent from controls. After 4 weeks, mean survival of 1.0% XP-fed shrimp (63.4 F 8.8%)remained higher than that of controls but the difference was not significant ( p = 0.07). Aninsufficient number of animals were available from the 0.5% XP-fed group to perform bacterialchallenges at this timepoint. Mean survival of control shrimp declined significantly over the 4 weeksof study (slope of linear regression p 0, p = 0.005, df = 11), but no decline was observed in animalsfed the 0.5% or 1.0% XP diets. After 4 weeks L. vannamei fed standard shrimp pellets, 0% XPcontrol, or 1.0% XP diets showed no significant differences in weight, suggesting that the changes indisease resistance did not correlate with changes in growth rate among the treatment groups. These
* Corresponding author. Tel.: +1-843-953-9171; fax: +1-843-953-9199. E-mail address: [email protected] (K.G. Burnett).
0044-8486/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.aquaculture.2003.09.003
J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
results indicate that dietary administration of Diamond V XP Yeast CultureR can protect shrimpagainst a decline in resistance to bacterial disease. D 2004 Elsevier B.V. All rights reserved.
Keywords: Litopenaeus vannamei; Diet; Yeast; Vibrio; Disease resistance
Productivity in aquaculture ponds is directly related to the growth rate and the survival of
animals stocked into the culture system. Losses attributed to bacterial, viral and fungaldiseases remain an important concern. Immunostimulants have been reported to increaseresistance to these infectious diseases in teleost fish and shellfish by enhancing the nonspecific immune system, the set of defenses directed against allpotentially invasive, disease-causing organisms. Two types of immunostimulants havereceived the most attention in shrimp aquaculture (1) fragments of bacterial cell walls, suchas lipopolysaccharide (LPS), and (2) beta-glucans from one of several fungal or algal species. Dietary exposure to Schizophyllum commune glucan significantly increasedresistance against white spot syndrome virus (WSSV) infection in Penaeus monodon post-larvae (15-day exposure) and juveniles (20-day exposure) and againstVibrio sp. NU-1 in Penaeus japonicus (7-day exposure,
Despite these favorable reports, immunostimulants have not yet been incorporated into
routine aquaculture of shrimp. cited several concerns of large-scale producers,including conflicting reports regarding the dose, timing and frequency of administration. reported that doses of peptidoglycan (PG) from the gram-positive bacterium Bifidobacterium thermophilum protected adult rainbow trout fromvibriosis but reduced the resistance of juvenile fish to infection with Vibrio sp. (1994) found that the immersion of P. monodon in high concentrations of glucan (>1 mg/ml) had adverse effects on shrimp, causing tissue damage and decreasing resistance toVibrio infection. In addition, reported that a diet of 0.1% glucan over 7weeks significantly reduced the ability of shrimp to clear live Vibrio harveyi from thehemolymph as compared to controls or to animals reared on diets containing 1% yeast,Saccahromyces cerevisiae.
With the exception of and relatively little
information is available regarding the performance of shrimp maintained on a dietsupplemented with yeast. In the present study, we tested whether a dried yeast culturesupplement (Diamond V XP Yeast CultureR, Diamond V Mills, Cedar Rapids, IA) mightbe used as an immunostimulant in the Pacific white shrimp, Litopenaeus vannamei.
To produce XP Yeast CultureR, aerobically fermented S. cerevisiae and its molasses-
based medium were mixed with cereal grains (ground yellow corn, hominy feed, corn
J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
gluten feed, wheat middlings and rye middlings) to form a slurry and fermented a secondtime under anaerobic conditions. After drying, the mixture was ground to produce thedesired final consistency of particle size. The grain carrier used for control diets consistedof the same cereal mixture in the same proportion as that used in the production of the XP.
Experimental diets were produced by combining commercial shrimp pellets (Rangen
35/2.5) with powdered XP and/or grain carrier and 50% gelatin (Knox) to produce threediets: (1) 1% XP, (2) 0.5% XP (containing 0.5% XP + 0.5% carrier), or (3) 0% Control XP(containing 1% carrier). Dry components were ground to a powder using a mortar andpestle. Gelatin (Knox), dissolved at 60 Ag/ml in boiling water, was cooled to 25 jC andmixed at 50% v/w with the powdered diet. The resulting paste was spread evenly onaluminum foil and cooled at 4 jC until solid. Pellets were then cut to approximately 0.5cm3 cubes and stored in sealed plastic bags at 4 jC for no longer than 7 days prior to use.
Post-larvae of L. vannamei (Kona High Health Stock, High Health Aquaculture, Kona,
HI) were grown to approximately 1 g juvenile stage animals at the Waddell MaricultureCenter, Bluffton, South Carolina (SC). These juveniles were transported to the GriceMarine Laboratory, Charleston, SC, where they were placed in recirculating seawater at30-ppt salinity and 23 – 26 jC throughout the experiments. Three days after transport,shrimp were placed on one of three experimental diets: 0% XP Control, 0.5% XP or 1%XP. Animals were fed ad lib one time each day. When shrimp had finished feeding (after 2to 3 h), uneaten food was removed from the tanks. Ammonia, pH and temperature of thewater were monitored daily and partial water exchanges were conducted as needed basedon these measurements. After 1, 2, 3 and 4 weeks on the experimental feeds, animals fromeach group were tested for disease resistance using a bacterial challenge model.
Each challenge test employed 21 shrimp that were injected intramuscularly in the third
abdominal segment with live Vibrio sp. 90-69B3 (kindly provided by D. Lightner and L. Mahone, University of Arizona). The 16S rRNA sequence of this strain places it in theVibrio parahaemolyticus/V. harveyi family (unpublished, Eric Stabb, University ofGeorgia). After 1, 2 and 4 weeks on the experimental diets, shrimp weighing from 0.5to 1.5 g (average approximately 1.0 g) were injected with 5 Al of 90-69B3 at aconcentration of 4 Â 107 cells/ml, based on an optical density (OD) value of 0.1 at 540nm being equal to 1 Â 108 bacteria/ml Larger animals (1.5 – 3.0 g,average approximately 2.0 g) that were challenged after 3 weeks on the test diets wereinjected with 10 Al of 90-69B3 at 4 Â 107 cells/ml, so that all animals in the study werechallenged with approximately 2 Â 105 bacteria per g body weight.
Following bacterial challenge, shrimp were held in a 19-l tank equipped with a
recirculating biological filter. Survival was monitored every 4 h for 48 h. Ammonia levelswere monitored and not allowed to exceed 1.0 mg/ml, although most readings were below0.25 mg/ml. Groups of 21 animals from each of the three diet groups were challengedsimultaneously using a single preparation of 90-69B3. Independent replicates with freshbacterial preparations were performed on days 8, 9, 10 (designated week 1), 15, 16, 17(designated week 2), 23, 24, 25 (designated week 3), 33 and twice on day 34 (designatedweek 4) with one exception. Animals from the 0.5% diet group were included in weeks 1,
J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
2 and 3 but not in week 4 challenges because there were not enough remaining animals ofappropriate size.
To compare the effects of diets on growth rate, 1 g juvenile stage animals with the same
history as those used in the disease resistance trials were placed in recirculating seawatertanks at 30 ppt and 23 – 26 jC. Groups of 25 – 27 shrimp were placed in 76-l tanksequipped with recirculating biofilters and maintained on standard shrimp pellets (Rangen35/2.5) or one of two experimental diets: 1% XP and 0% XP Control. Animals were fed adlib daily and tanks maintained as described above. Individual shrimp were weighed eachweek for 4 weeks.
A two-way Model I ANOVA was used to distinguish the effects of diet and duration of
feeding on 48-h survival and on growth over the 4 weeks of study, while one-wayANOVAs were performed on 48-h survival data from each week. Post hoc pairwisemultiple comparisons were conducted on significant effects using the Holm – Sidakmethod. Individual regressions were run on survival data within each diet group to assesschanges over the feeding period. For all statistical tests, a p value of 0.05 was used todetermine significance.
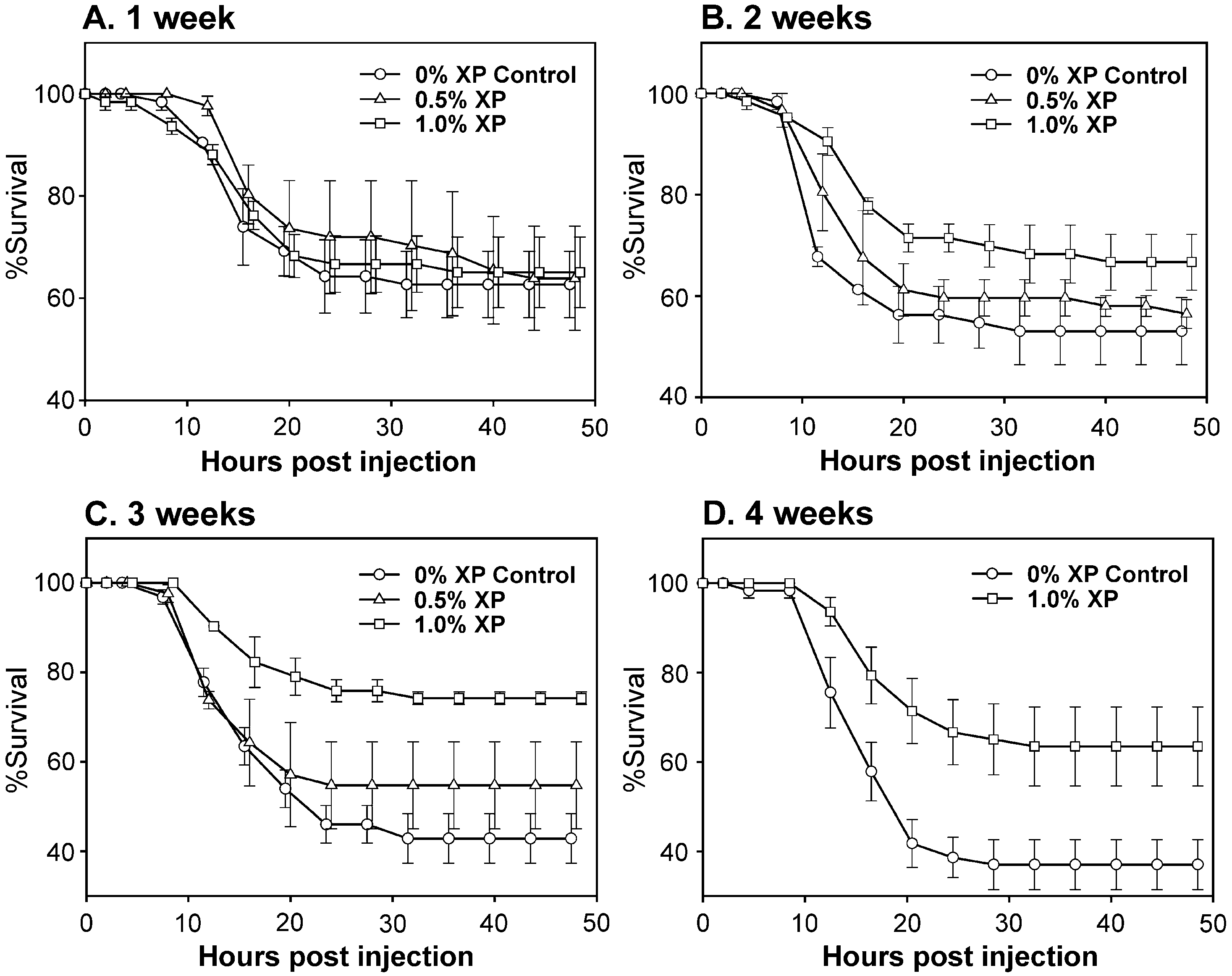
For animals on all diets, mortalities following bacterial challenge began to occur at 8- to
12-h post injection and ceased by 48-h post injection Two-way ANOVA showed asignificant effect of diet ( p = 0.003, df = 31) but not duration of feeding ( p = 0.193, df = 31)on 48-h survival. Post hoc pairwise multiple comparisons on the effect of diet showedsignificant differences in survival between animals on 1% XP and 0.5%XP ( p = 0.048) andbetween 1% XP and 0% XP Control diets ( p < 0.001).
After 1 or 2 weeks on the test diets, there were no significant differences in mean
survival at 48 h following bacterial challenge. After 3 weeks, mean survival of L. vannamei fed 1.0% XP was significantly higher than that of control shrimp (one-way ANOVA; p = 0.029, df = 7). Survivals after 48 h for 1% XP-fed shrimp in thethree replicate trials were 71%, 75% and 76% (mean 74.2 F 1.4% S.E.) as compared to33%, 42% and 52% (mean 42.9 F 5.5% S.E.) for controls. There was no significantdifference in 48-h survival between shrimp fed 0.5% XP and 0% XP Control diets. After 4weeks 48-h survivals of 1.0% XP-fed L. vannamei (52, 57, 81%) were higher thanthose of controls (29%, 35%, 48%), but the difference was not significant (one-wayANOVA; p = 0.065, df = 5). An insufficient number of animals from the 0.5% XP dietgroup were available for testing after 4 weeks. Within the 0.5% and 1.0% XP diet groups,48-h survival did not change significantly over the 4 weeks of the feeding trial (slope of linear regression p 0, p = 0.438, df = 7 and p = 0.922, df = 11, respectively). By
J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
Fig. 1. Survival time course of L. vannamei fed one of three test diets and challenged with Vibrio sp. 90-69B3. Mean survivals F standard error (S.E.) (n = 3, except C. 0.5% XP, n = 2) are given at 4-h intervals over 48 hfollowing bacterial challenge for each diet group on (A) week 1, (B) week 2, (C) week 3, (D) week 4.
Fig. 2. Summary of 48-h survivals of L. vannamei after injection with Vibrio sp. 90-69B3. Values are meansurvivals F S.E. (n = 3, except 0.5% at 3 weeks n = 2). (*) L. vannamei fed 1.0% XP had a significantly higheroverall survival than control (0% XP) shrimp after 3 weeks of feeding.
J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
Fig. 3. Growth of L. vannamei (n = 25 – 27) fed daily with standard shrimp pellets, 0% XP diet, or 1.0% XP dietfor 4 weeks. Shrimp were weighed each week and mean weights F S.E. are presented. There were no significantdifferences among the weights of the three diet groups at any timepoint tested.
comparison, the survival of control shrimp decreased over the 4-week trial (slope oflinear regression p = 0.005, df = 11). There were no significant differences in the meanweights of animals among the three diet groups that were challenged in each week’s trials,nor were there any significant differences among the average weights of animals used fromweek to week, with the exception of week 3 as described in Materials and methods (datanot shown). Furthermore, there were no significant differences in the growth of animalsmaintained on 0% XP Control, 1% XP or the standard shrimp pellet diet
The level of survival in response to bacterial challenge in L. vannamei remained
constant in animals fed the 1% XP supplement, while survival declined in control shrimpfed only the grain-based carrier. These results are consistent with those of and who observed greater viability in P. monodon fed a relatively loworal dose (0.2%) of beta-glucan from S. commune over 30 or 40 days. Similarly, combinedimmersion and dietary exposure to h-1,3-1,6-glucan from cell walls of the yeast S. cerevisiae for 51 days enhanced resistance of juvenile P. monodon to WSSV infectionIn contrast, the potentially negative effects associated with long-termuse of immunostimulants have led otherinvestigators to recommend their periodic administration. fed PG to P. monodon at 0.2% of diet for seven consecutive days alternated with 7 days without theimmunostimulant. These workers reported improved resistance to Vibrio penaeicida at 65and 95 days in PG-fed animals as compared to controls.
The present study did not address the mechanism by which the dried yeast culture
afforded protection. It is possible that the yeast-supplemented diet provided more optimal
J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
nutrition than the control diet, but this was not reflected in the growth rates. The protectiveeffect of the yeast supplement might also be attributed to its glucan content. Glucans arereported to enhance disease resistance by stimulating nonspecific components of theimmune system or by improving processing and presentation of antigens during specificadaptive immune responses. For example, the protective effects of beta-glucan incrustaceans have been associated with activation of the prophenoloxidase system inducingantimicrobial activity in plasma and enhancing phagocytosis, cell adhesion and superoxideproduction in hemocytes Chang et al., 2000; Campa-Cordova et al., 2002).
Shrimp fed the control diet exhibited a decline in resistance to bacterial challenge. The
reasons for this decline are not clear but may be attributed to one or more of the followinginterdependent factors. First, juvenile shrimp used in these studies came from closedrecirculating production raceways that supported a dense microbial flora. These exposuresmight enhance the basal activity of the immune system in crustaceans. Once transferred tolaboratory tanks with filtered seawater, test animals received much lower environmentaland dietary exposures to bacteria, fungi or algae. With reduced exposure to activationsignals from environmental or dietary microbes, the immune system of crustaceans mayenter into a lower state of readiness. This phenomenon, called immunoquiesence, has beendocumented in the purple sea urchin Strongylocentrotus purpuratus maintained for 18months in UV-sterilized, artificial seawater on a diet of dried kelp 2000). The yeast supplement might replace the absent microbial flora to maintain theactivated state of immunological protection in the shrimp. Second, the laboratoryconditions under which this study was conducted might have been stressful, leading toimmunosuppression by any one of a number of mechanisms that have been suggested inthe scientific literature. In this case, the yeast supplement may have protected against thenegative effects of stress on the immune system. Finally, microbial flora in productionraceways or ponds might normally serve as a nutritional supplement to the basal diet. Oncetransferred to filtered seawater, shrimp on the basal diet alone might have lost thatcomponent of dietary nutrition leading to a general decline in animal health. The yeastadditive may have replaced the missing component of dietary nutrition. Regardless of theunderlying reason(s) for its occurrence, this temporal decline in disease resistance amongcontrol shrimp maintained under laboratory conditions should be considered in evaluatingthe efficacy of immunostimulants.
Further laboratory studies as well as field trials are required to confirm the efficacy of
Diamond V XP Yeast CultureR. Caution always must be taken in translating laboratoryresults to farm application. Nonetheless, it is clear that feed supplements that are readilyavailable in large-scale quantities and have the capacity to adapt to high salt concentrationsdeserve additional study as immunostimulants in aquaculture.
This study was supported by Diamond V Mills, Cedar Rapids, IA. The authors thank
Dr. Craig Browdy and The Waddell Mariculture Center, SC Department of NaturalResources, for providing L. vannamei used in these experiments. This manuscript isContribution No. 234 from the Grice Marine Laboratory.
J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
Adler, L., 1994. Response of the yeast Saccharomyces cerevisiae to salt stress. 3rd International Marine Bio-
technology Conference, Program Abstracts. International Advisory Comm. of the Int. Marine BiotechnologyConference. Tromsoe, Norway, p. 46.
Campa-Cordova, A.I., Hernandez-Saaverdra, N.Y., DePhilippis, R., Ascencio, R., 2002. Generation of super-
oxide anion and SOD activity in haemocytes and muscle of American white shrimp (Litopeaneus vannamei)as a response to h-glucan and sulphated polysaccharide. Fish Shellfish Immunol. 353 – 366.
Chang, C.F., Su, M.S., Chen, H.Y., Lo, C.F., Kou, G.H., Liao, I.C., 1999. Effect of dietary beta-1,3-glucan on
resistance to white spot syndrome virus (WSSV) in postlarval and juvenile Penaeus monodon. Dis. Aquat. Org. 36, 163 – 168.
Chang, C.-F., Chen, H.-Y., Su, M.-S., Liao, I.-C., 2000. Immunomodulation by dietary h-1,3 glucan in the
brooders of the black tiger shrimp Penaeus monodon. Fish Shellfish Immunol. 10, 505 – 514.
Gross, P.S., Al-Sharif, W.Z., Clow, L.A., Smith, L.C., 1999. Echinoderm immunity and the evolution of the
complement system. Dev. Comp. Immunol. 23, 439 – 442.
Gross, P.S., Clow, L.A., Smith, L.C., 2000. SpC3, the complement homologue from the purple sea urchin,
Strongylocentrotus purpuratus, is expressed in two subpopulations of the phagocytic coelomocytes. Immu-nogenetics 51, 1034 – 1044.
Itami, T., Takahashi, Y., Tsuchihira, E., Igusa, H., Konda, M., 1994. Enhancement of disease resistance of
kuruma prawn Penaeus japonicus and increase in phagocytic activity of prawn hemocytes after oraladministration of h-1,3-glucan (Schizophyllan). In: Chou, L.M., Munro, A.D., Lam, J.J., Chen, T.W.,Cheong, L.K.K., Ding, J.K., Hooi, K.K., Khoo, H.W., Phang, V.P.E., Shim, K.F., Tan, C.H. (Eds.), TheThird Asian Fisheries Forum. Asian Fisheries Society, Manila, Philippines, pp. 375 – 378.
Itami, T., Asano, M., Tokushige, K., Kubono, K., Nakagawa, A., Takeno, N., Nishimura, H., Maeda, M.,
Kondo, M., Takahashi, Y., 1998. Enhancement of disease resistance of kuruma shrimp, Penaeus japonicus,after oral administration of peptidoglyan derived from Bifidobacterium thermophilum. Aquaculture 164,277 – 288.
Matsuo, K., Miyazono, I., 1993. The influence of long-term administration of peptidoglycan on disease resistance
and growth of juvenile rainbow trout. Nippon Suisan Gakkaishi 59, 1377 – 1379.
Mikulski, C.M., Burnett, L.E., Burnett, K.G., 2000. The effects of hypercapnic hypoxia on the survival of shrimp
challenged with Vibrio parahaemolyticus. J. Shellfish Res. 19, 301 – 311.
Raa, J., 1996. The use of immunostimulatory substances in fish and shellfish farming. Reviews Fish. Sci. 4,
Sakai, M., 1999. Current research status of fish immunostimulants. Aquaculture 172, 63 – 92. Scholz, U., Diaz, G.G., Ricque, D., Cruz Suarez, L.E.C., Albores, F.V., Latchford, J., 1999. Enhancement of
vibriosis resistance in juvenile Penaeus vannamei by supplementation of diets with different yeast products. Aquaculture 176, 271 – 283.
Song, Y.L., Hsieh, Y.T., 1994. Immunostimulation of tiger shrimp (Penaeus monodon) hemocytes for generation
of microbicidal substances: analysis of reactive oxygen species. Dev. Comp. Immunol. 18, 102 – 209.
Song, Y.L., Liu, J.J., Chan, L.C., Sung, H.H., 1997. Glucan-induced disease resistance in tiger shrimp (Penaeus
monodon). Fish Vaccinol.: Dev. Biol. Stand. 90, 413 – 421.
Su, M.S., Liu, K.F., Chang, C.F., Liao, I.C., 1995. Enhancement of grass prawn Penaeus monodon postlarvae
viability by beta-1,3-glucan from Schizophyllum commune. J. Taiwan Fish. Res. 3, 125 – 132 (in Chinese withEnglish abstract).
Sung, H.H., Kou, G.H., Song, Y.L., 1994. Vibriosis resistance induced by glucan treatment in tiger shrimp
(Penaeus monodon). Fish Pathol. 29, 11 – 17.
Sung, H.H., Yang, Y.L., Song, Y.L., 1996. Enhancement of microbicidal activity in the tiger shrimp (Penaeus
monodon) via immunostimulation. J. Crustac. Biol. 16, 278 – 284.
FIORENZO CONTI 1. PERSONAL HISTORY Department of Experimental & Clinical Medicine Section of Neuroscience & Cell Biology Università Politecnica delle Marche Via Tronto 10/A, Torrette di Ancona I-60020 Ancona (Italy) Phone 0039 071 220 6056 Faz [email protected] www.fiorenzocontigroup.it Birth Date and Place August 13, 1955 Villerupt (France) Current position
Ophthalmological Society of South Africa Preliminary Programme 24 – 27 March 2011 CONTENTS OPHTHALMIC PROGRAMME THURSDAY, 24 MARCH 2011 MEETINGS : Council Meeting: College of Ophthalmologists REFRESHMENTS FOR ALL DELEGATES SESSION 1 Retina & Oncology Chair: Dr Karin Lecuona 10:40 Whats' new in malignant intraocular tumors? Prof Jerry Shields, USA (


 Disease resistance of Pacific white shrimp,
Litopenaeus vannamei, following the dietary
administration of a yeast culture food supplement
Joseph E. Burgents, Karen G. Burnett*, Louis E. Burnett
Grice Marine Laboratory, College of Charleston, 205 Fort Johnson Road, Charleston, SC 29412, USA
Received 19 February 2003; received in revised form 20 August 2003; accepted 2 September 2003
A yeast culture feed supplement (Diamond V XP Yeast CultureR, Diamond V Mills, Cedar
Rapids, Iowa [IA]) was assessed for its impact on disease resistance in the Pacific white shrimp,Litopenaeus vannamei. Animals were fed a standard shrimp pellet diet supplemented with 0%(control with 1% grain carrier), 0.5% (with 0.5% carrier), or 1.0% XP daily for 4 weeks. To assessresistance to bacterial disease, at 1-week intervals 21 shrimp (0.5 – 2.5 g) from each test diet wereinjected intramuscularly with an LD50 dose (2.0 Â 105/g body weight) of a gram-negative shrimppathogen, Vibrio sp. 90-69B3. Survival was monitored every 4 h for 48-h post injection. Each week,three independent bacterial challenges were performed for each diet and the results expressed as themean percent survival F standard error (S.E.). A two-way analysis of variance (ANOVA) showed asignificant effect of diet ( p = 0.003, df = 31), but not duration of feeding, on survival. A one-wayANOVA showed no differences among the treatment groups after 1 or 2 weeks. After 3 weeks, themean survival of 1% XP-fed shrimp (74.2 F 1.4%) was significantly higher than that of controls(42.9 F 5.5%), while mean survival of shrimp fed 0.5% XP (54.8 F 11.9%) was not significantlydifferent from controls. After 4 weeks, mean survival of 1.0% XP-fed shrimp (63.4 F 8.8%)remained higher than that of controls but the difference was not significant ( p = 0.07). Aninsufficient number of animals were available from the 0.5% XP-fed group to perform bacterialchallenges at this timepoint. Mean survival of control shrimp declined significantly over the 4 weeksof study (slope of linear regression p 0, p = 0.005, df = 11), but no decline was observed in animalsfed the 0.5% or 1.0% XP diets. After 4 weeks L. vannamei fed standard shrimp pellets, 0% XPcontrol, or 1.0% XP diets showed no significant differences in weight, suggesting that the changes indisease resistance did not correlate with changes in growth rate among the treatment groups. These
* Corresponding author. Tel.: +1-843-953-9171; fax: +1-843-953-9199.
Disease resistance of Pacific white shrimp,
Litopenaeus vannamei, following the dietary
administration of a yeast culture food supplement
Joseph E. Burgents, Karen G. Burnett*, Louis E. Burnett
Grice Marine Laboratory, College of Charleston, 205 Fort Johnson Road, Charleston, SC 29412, USA
Received 19 February 2003; received in revised form 20 August 2003; accepted 2 September 2003
A yeast culture feed supplement (Diamond V XP Yeast CultureR, Diamond V Mills, Cedar
Rapids, Iowa [IA]) was assessed for its impact on disease resistance in the Pacific white shrimp,Litopenaeus vannamei. Animals were fed a standard shrimp pellet diet supplemented with 0%(control with 1% grain carrier), 0.5% (with 0.5% carrier), or 1.0% XP daily for 4 weeks. To assessresistance to bacterial disease, at 1-week intervals 21 shrimp (0.5 – 2.5 g) from each test diet wereinjected intramuscularly with an LD50 dose (2.0 Â 105/g body weight) of a gram-negative shrimppathogen, Vibrio sp. 90-69B3. Survival was monitored every 4 h for 48-h post injection. Each week,three independent bacterial challenges were performed for each diet and the results expressed as themean percent survival F standard error (S.E.). A two-way analysis of variance (ANOVA) showed asignificant effect of diet ( p = 0.003, df = 31), but not duration of feeding, on survival. A one-wayANOVA showed no differences among the treatment groups after 1 or 2 weeks. After 3 weeks, themean survival of 1% XP-fed shrimp (74.2 F 1.4%) was significantly higher than that of controls(42.9 F 5.5%), while mean survival of shrimp fed 0.5% XP (54.8 F 11.9%) was not significantlydifferent from controls. After 4 weeks, mean survival of 1.0% XP-fed shrimp (63.4 F 8.8%)remained higher than that of controls but the difference was not significant ( p = 0.07). Aninsufficient number of animals were available from the 0.5% XP-fed group to perform bacterialchallenges at this timepoint. Mean survival of control shrimp declined significantly over the 4 weeksof study (slope of linear regression p 0, p = 0.005, df = 11), but no decline was observed in animalsfed the 0.5% or 1.0% XP diets. After 4 weeks L. vannamei fed standard shrimp pellets, 0% XPcontrol, or 1.0% XP diets showed no significant differences in weight, suggesting that the changes indisease resistance did not correlate with changes in growth rate among the treatment groups. These
* Corresponding author. Tel.: +1-843-953-9171; fax: +1-843-953-9199.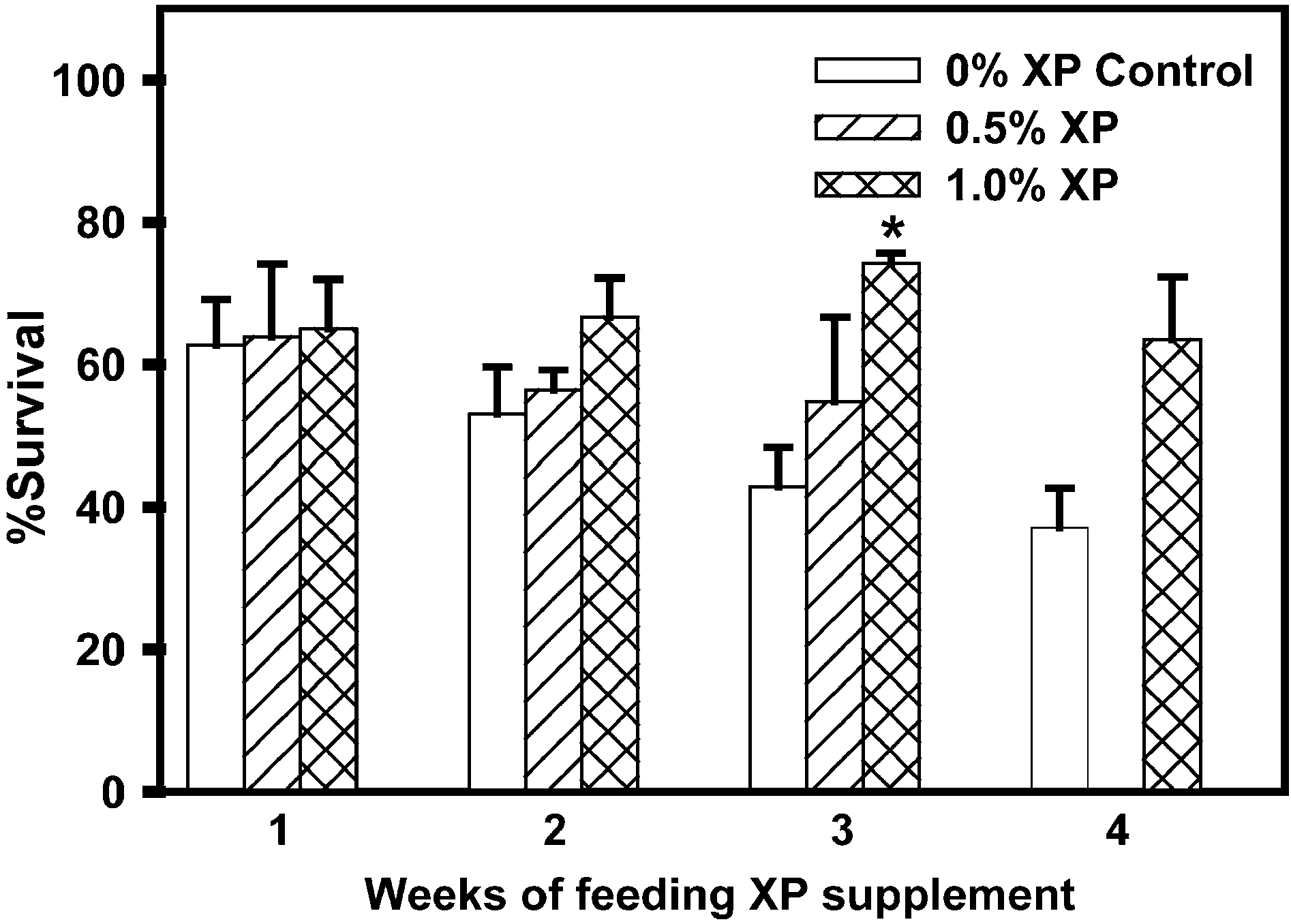
 J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
Fig. 1. Survival time course of L. vannamei fed one of three test diets and challenged with Vibrio sp. 90-69B3.
J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
Fig. 1. Survival time course of L. vannamei fed one of three test diets and challenged with Vibrio sp. 90-69B3.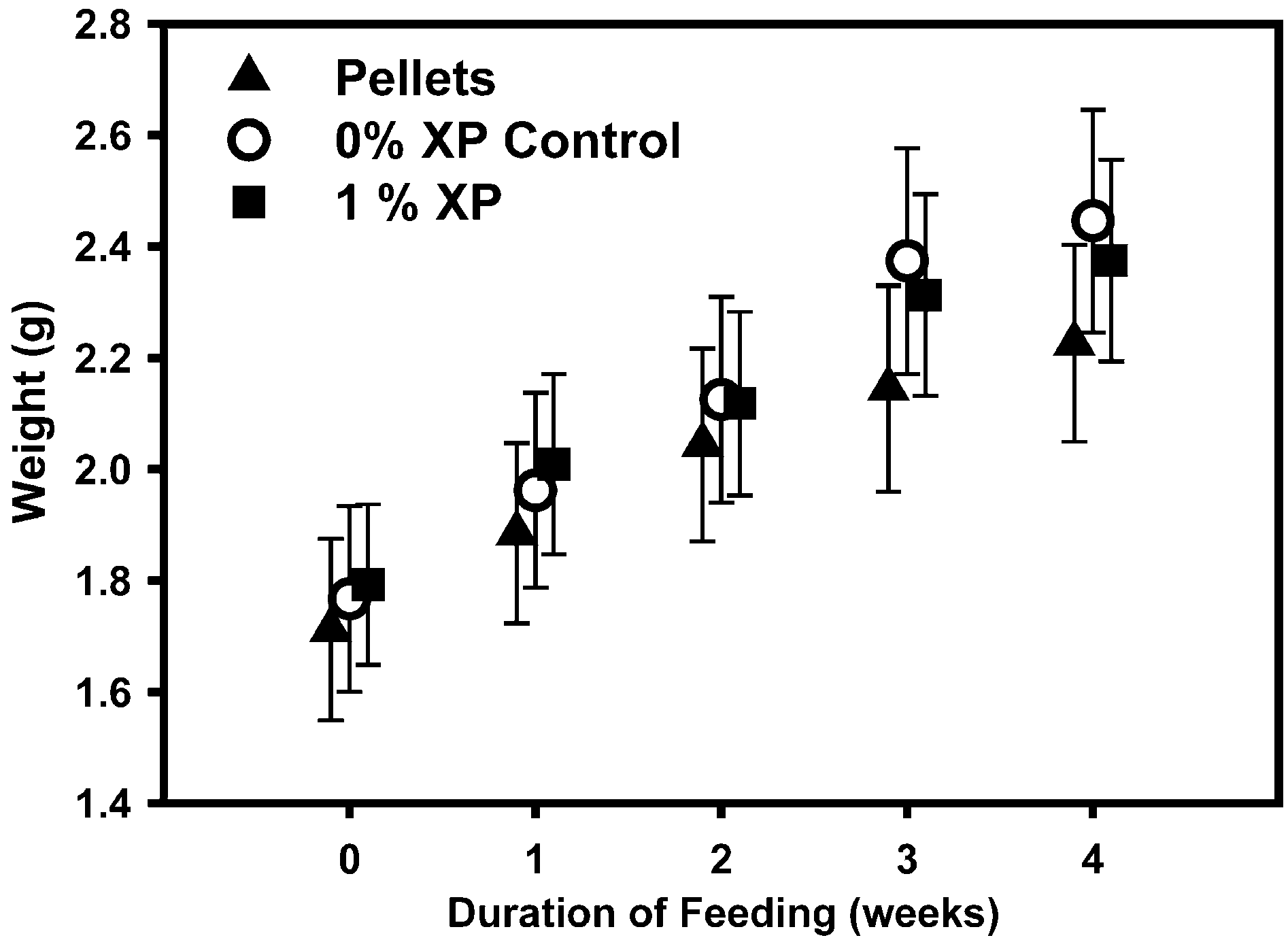 J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
Fig. 3. Growth of L. vannamei (n = 25 – 27) fed daily with standard shrimp pellets, 0% XP diet, or 1.0% XP dietfor 4 weeks. Shrimp were weighed each week and mean weights F S.E. are presented. There were no significantdifferences among the weights of the three diet groups at any timepoint tested.
J.E. Burgents et al. / Aquaculture 231 (2004) 1–8
Fig. 3. Growth of L. vannamei (n = 25 – 27) fed daily with standard shrimp pellets, 0% XP diet, or 1.0% XP dietfor 4 weeks. Shrimp were weighed each week and mean weights F S.E. are presented. There were no significantdifferences among the weights of the three diet groups at any timepoint tested.